Az inaktivált SARS-CoV-2 biodisztribuciója és sejtes internalizálása az egerekben



1
Pharmacoidea Kft., 6726 Szeged
2
Boeckeler Instruments, Inc., Tucson, AZ 85714, Amerikai Egyesült Államok
3
Delong Instruments a.s., 612 00 Brno, Csehország
4
Biofizikai Intézet, Biológiai Kutatóközpont, 6726 Szeged, Magyarország
5
Szegedi Tudományegyetem, Általános Orvostudományi Kar, MÁSZFok, 6720 Szeged
6
Szegedi Tudományegyetem, Szent-Györgyi Albert Klinikai Központ, MÁSZTAN, 6720 Szeged, MTA 6720
7
Proteomikai Kutató Laboratórium, Biológiai Kutatóközpont, 6726 Szeged, 6726
8
Biokémiai Intézet, Biológiai Kutatóközpont, 6726 Szeged
*
Szerző, akinek a levelezést kell címezni.
Akadémiai szerkesztők: Malgorzata Kloc és Jacek Z. Kubiak
Int. J. Mol. Sci. 2022, 23(14), 7609; https://doi.org/10.3390/ijms23147609
Beérkezett: 2022. június 1. / Módosítva: 2022. július 4. / Elfogadva: 2022. július 7. / Közzétéve: 2022. július 9.
Az inaktivált SARS-CoV-2 biodisztribuciója és sejtes internalizálása vad típusú egerekben
1
Pharmacoidea Kft., 6726 Szeged
2
Boeckeler Instruments, Inc., Tucson, AZ 85714, Amerikai Egyesült Államok
3
Delong Instruments a.s., 612 00 Brno, Csehország
4
Biofizikai Intézet, Biológiai Kutatóközpont, 6726 Szeged, Magyarország
5
Szegedi Tudományegyetem, Általános Orvostudományi Kar, MÁSZFok, 6720 Szeged
6
Szegedi Tudományegyetem, Szent-Györgyi Albert Klinikai Központ, MÁSZTAN, 6720 Szeged, MTA 6720
7
Proteomikai Kutató Laboratórium, Biológiai Kutatóközpont, 6726 Szeged, 6726
8
Biokémiai Intézet, Biológiai Kutatóközpont, 6726 Szeged
*
Szerző, akinek a levelezést kell címezni.
Akadémiai szerkesztők: Malgorzata Kloc és Jacek Z. Kubiak
Int. J. Mol. Sci. 2022, 23(14), 7609; https://doi.org/10.3390/ijms23147609
Beérkezett: 2022. június 1. / Módosítva: 2022. július 4. / Elfogadva: 2022. július 7. / Közzétéve: 2022. július 9.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Az azonosított SARS-CoV-2 receptorok növekvő listája ellenére a humán angiotenzin-konvertáló 2 enzimet (ACE2) továbbra is a SARS-CoV-2 internalizációt közvetítő fő sejtbelépési receptornak tekintik. Beszámoltak arról, hogy a vadon élő egerek, mint a Muridae család más rágcsálófajai, nem fertőződhetnek meg SARS-CoV-2-vel az ACE2 receptoraik különbségei miatt. Másrészt a SARS-CoV-2 tüskefehérjéjének, a PRRAR-nak a konszenzusos heparinkötő motívuma lehetővé teszi a rágcsálók heparán-szulfát proteoglikánjaihoz (HSMG-k) való kötődését, beleértve a szindekánokat is, egy transzmembrán HSPG családot, amely jól megalapozott szerepet játszik a klatrin- és caveolin-független endocitózisban. Mivel az emlős szindekánok viszonylag megőrzött szerkezettel rendelkeznek, elemeztük az inaktivált SARS-CoV-2 részecskék sejtfelvételét in vitro és in vivo egerek modelljeiben. A sejtes vizsgálatok hatékony felvételt mutattak ki a purin sejtvonalakon, amelyeken kialakult a szindekán-4 expresszió. Intravénás beadás után az inaktivált SARS-CoV-2-t több szerv vette fel in vivo, és az agyban is kimutatható volt. A különböző szövetek által internalizált inaktivált SARS-CoV-2 megemelte a szöveti TNF-α szinteket, különösen a szívben, tükrözve a gyulladás kialakulását. Az in vitro és in vivo egerek modelljein végzett vizsgálataink így fényt derítenek a SARS-CoV-2 internalizáció ismeretlen részleteire, és segítenek kiszélesíteni a SARS-CoV-2 molekuláris kölcsönhatásainak megértését.
Kulcsszavak: SARS-CoV-2; celluláris felvétel; heparán-szulfát proteoglikánok; szindekánok; egér
1. Bevezetés
A koronavírus 2 (SARS-CoV-2) súlyos akut légzőszervi szindróma félelmetes kihívások elé állítja a klinikusokat és a kutatókat világszerte [1,2]. A SARS-CoV-2 fertőzés molekuláris hátterének növekvő részletei lehetővé tették a hatékony vakcinák és új vírusellenes szerek kifejlesztését [3,4]. Az azonosított SARS-CoV-2 receptorok növekvő listája ellenére a humán angiotenzin-konvertáló 2 enzimet (ACE2) még mindig a SARS-CoV-2 fertőzés során a fő sejtbelépési receptornak tekintik [5,6]. A legújabb tanulmányok azonban azt mutatják, hogy a SARS-CoV-2 ACE2-független sejtes bejegyzése [7,8,9]. Az ACE2 receptorok különbségei miatt a vadon élő típusú (WT) egerek állítólag nem fertőződhetnek meg SARS-CoV-2-vel [10,11]. Másrészt a SARS-CoV-2 tüskefehérjéjében, a PRRAR-ban található evolúciósan megőrzött heparinkötő motívum lehetővé teszi az emlős heparán-szulfát proteoglikánokhoz (HSMG-k), más jól bevált SARS-CoV-2 receptorokhoz [12,13,14,15,16,17] való kötődést.
Korábban azt vizsgáltuk, hogy a syndekánok (SFC-k), egy evolúciósan konzervált transzmembrán HSPG család, amely jól megalapozott szerepet tölt be a klathrin- és caveolin-független endocitózisban, hogyan járul hozzá a SARS-CoV-2 sejtes internalizációjához [18,19,20,21,22,23]. Eredményeink szerint az SDC-k közvetítik a SARS-CoV-2 sejtes belépését azáltal, hogy csatolják a heparinkötő motívumot (PRRAR) tartalmazó S1 alegységet. Az SFC-k közül a syndecan-4 (SDC4), a tüdőben dúsított izoforma közvetítette a SARS-CoV-2 sejtfelvételét és Delta variánsát a leghatékonyabban [18,24]. A heparinnal vagy heparinkötő peptidekkel való kompetitív gátlás csökkentette a SARS-CoV-2 sejtbejutását, bizonyítva a heparán-szulfáthoz (HS) való kötődés fontosságát.
A glikozaminoglikánok (GAG-ok) családjába tartozó szulfátosított poliszacharid, a HS, a HS, a glikozaminoglikánok (GAG-ok) családjába tartozó szulfátos poliszacharid mindenütt eloszlik az állati sejtek felszínén és az extracelluláris mátrixban [25,26]. A fehérje-GAG kölcsönhatások kiemelkedő szerepet játszanak a sejt-sejt kölcsönhatásokban, a sejtek növekedésében és a vírusfertőzésekben [27,28,29]. Az XBBXBX vagy XBBBXXBX konszenzusos heparin/HS-kötő motívumok (B az arginin, lizin vagy hisztidin bázikus aminosavak, az X pedig az alifás/aromás aminosavak egyik tartománya) fehérjéket biztosítanak a HS-hez való kötődéshez [30,31]. Megőrzött struktúráik miatt a purin SFC-k nagyon azonosak emberi társaikkal. Amellett, hogy ektodomainjeik 70%-ban azonosak, minden feltételezett glikozaminoglikán kötődési hely azonos. Ezenkívül a transzmembrán domén 96%-ban azonos, míg a citoplazmatikus domén 100%-ban azonos, beleértve három azonosan elhelyezkedő tirozinmaradékot [32]. Az emlős HSSPG-k szulfatációs mintázatának finom különbségei ellenére a konszenzusos HS-kötő motívumok, mint amilyenek a SARS-CoV-2 tüskéjében jelen vannak, lehetővé teszik a heparinkötő fehérjék emlősök HSPG-hez, köztük SFC-khez való kötődését [17,25,33]. Az SDC által közvetített endocitózis során a ligandumok kötődése kiváltja az SFC-k klaszterezését és serkenti az SDC–ligandum komplex internalizációját [20,34,35].
Tekintettel a SARS-CoV-2 HS-2 tüskék HS-kötő motívumának affinitására a HS-hez és a HSSPG-k megalapozott szerepére a nem beklasszikus endocitózisban, úgy döntöttünk, hogy elemezzük az inaktivált SARS-CoV-2 részecskék sejtfelvételét in vitro és in vivo egerek modelljeiben. A sejt- és állatkísérletek lehetővé tették a SARS-CoV-2 sejtbelépés fókuszált elemzését, és feltárták a SARS-CoV-2 internalizáció új aspektusait.
Az inaktivált SARS-CoV-2 biodisztribuciója és sejtes internalizálása vad típusú egerekben
1
Pharmacoidea Kft., 6726 Szeged
2
Boeckeler Instruments, Inc., Tucson, AZ 85714, Amerikai Egyesült Államok
3
Delong Instruments a.s., 612 00 Brno, Csehország
4
Biofizikai Intézet, Biológiai Kutatóközpont, 6726 Szeged, Magyarország
5
Szegedi Tudományegyetem, Általános Orvostudományi Kar, MÁSZFok, 6720 Szeged
6
Szegedi Tudományegyetem, Szent-Györgyi Albert Klinikai Központ, MÁSZTAN, 6720 Szeged, MTA 6720
7
Proteomikai Kutató Laboratórium, Biológiai Kutatóközpont, 6726 Szeged, 6726
8
Biokémiai Intézet, Biológiai Kutatóközpont, 6726 Szeged
*
Szerző, akinek a levelezést kell címezni.
Akadémiai szerkesztők: Malgorzata Kloc és Jacek Z. Kubiak
Int. J. Mol. Sci. 2022, 23(14), 7609; https://doi.org/10.3390/ijms23147609
Beérkezett: 2022. június 1. / Módosítva: 2022. július 4. / Elfogadva: 2022. július 7. / Közzétéve: 2022. július 9.
Az azonosított SARS-CoV-2 receptorok növekvő listája ellenére a humán angiotenzin-konvertáló 2 enzimet (ACE2) továbbra is a SARS-CoV-2 internalizációt közvetítő fő sejtbelépési receptornak tekintik. Beszámoltak arról, hogy a vadon élő egerek, mint a Muridae család más rágcsálófajai, nem fertőződhetnek meg SARS-CoV-2-vel az ACE2 receptoraik különbségei miatt. Másrészt a SARS-CoV-2 tüskefehérjéjének, a PRRAR-nak a konszenzusos heparinkötő motívuma lehetővé teszi a rágcsálók heparán-szulfát proteoglikánjaihoz (HSMG-k) való kötődését, beleértve a szindekánokat is, egy transzmembrán HSPG családot, amely jól megalapozott szerepet játszik a klatrin- és caveolin-független endocitózisban. Mivel az emlős szindekánok viszonylag megőrzött szerkezettel rendelkeznek, elemeztük az inaktivált SARS-CoV-2 részecskék sejtfelvételét in vitro és in vivo egerek modelljeiben. A sejtes vizsgálatok hatékony felvételt mutattak ki a purin sejtvonalakon, amelyeken kialakult a szindekán-4 expresszió. Intravénás beadás után az inaktivált SARS-CoV-2-t több szerv vette fel in vivo, és az agyban is kimutatható volt. A különböző szövetek által internalizált inaktivált SARS-CoV-2 megemelte a szöveti TNF-α szinteket, különösen a szívben, tükrözve a gyulladás kialakulását. Az in vitro és in vivo egerek modelljein végzett vizsgálataink így fényt derítenek a SARS-CoV-2 internalizáció ismeretlen részleteire, és segítenek kiszélesíteni a SARS-CoV-2 molekuláris kölcsönhatásainak megértését.
Kulcsszavak: SARS-CoV-2; celluláris felvétel; heparán-szulfát proteoglikánok; szindekánok; egér
1. Bevezetés
A koronavírus 2 (SARS-CoV-2) súlyos akut légzőszervi szindróma félelmetes kihívások elé állítja a klinikusokat és a kutatókat világszerte [1,2]. A SARS-CoV-2 fertőzés molekuláris hátterének növekvő részletei lehetővé tették a hatékony vakcinák és új vírusellenes szerek kifejlesztését [3,4]. Az azonosított SARS-CoV-2 receptorok növekvő listája ellenére a humán angiotenzin-konvertáló 2 enzimet (ACE2) még mindig a SARS-CoV-2 fertőzés során a fő sejtbelépési receptornak tekintik [5,6]. A legújabb tanulmányok azonban azt mutatják, hogy a SARS-CoV-2 ACE2-független sejtes bejegyzése [7,8,9]. Az ACE2 receptorok különbségei miatt a vadon élő típusú (WT) egerek állítólag nem fertőződhetnek meg SARS-CoV-2-vel [10,11]. Másrészt a SARS-CoV-2 tüskefehérjéjében, a PRRAR-ban található evolúciósan megőrzött heparinkötő motívum lehetővé teszi az emlős heparán-szulfát proteoglikánokhoz (HSMG-k), más jól bevált SARS-CoV-2 receptorokhoz [12,13,14,15,16,17] való kötődést.
Korábban azt vizsgáltuk, hogy a syndekánok (SFC-k), egy evolúciósan konzervált transzmembrán HSPG család, amely jól megalapozott szerepet tölt be a klathrin- és caveolin-független endocitózisban, hogyan járul hozzá a SARS-CoV-2 sejtes internalizációjához [18,19,20,21,22,23]. Eredményeink szerint az SDC-k közvetítik a SARS-CoV-2 sejtes belépését azáltal, hogy csatolják a heparinkötő motívumot (PRRAR) tartalmazó S1 alegységet. Az SFC-k közül a syndecan-4 (SDC4), a tüdőben dúsított izoforma közvetítette a SARS-CoV-2 sejtfelvételét és Delta variánsát a leghatékonyabban [18,24]. A heparinnal vagy heparinkötő peptidekkel való kompetitív gátlás csökkentette a SARS-CoV-2 sejtbejutását, bizonyítva a heparán-szulfáthoz (HS) való kötődés fontosságát.
A glikozaminoglikánok (GAG-ok) családjába tartozó szulfátosított poliszacharid, a HS, a HS, a glikozaminoglikánok (GAG-ok) családjába tartozó szulfátos poliszacharid mindenütt eloszlik az állati sejtek felszínén és az extracelluláris mátrixban [25,26]. A fehérje-GAG kölcsönhatások kiemelkedő szerepet játszanak a sejt-sejt kölcsönhatásokban, a sejtek növekedésében és a vírusfertőzésekben [27,28,29]. Az XBBXBX vagy XBBBXXBX konszenzusos heparin/HS-kötő motívumok (B az arginin, lizin vagy hisztidin bázikus aminosavak, az X pedig az alifás/aromás aminosavak egyik tartománya) fehérjéket biztosítanak a HS-hez való kötődéshez [30,31]. Megőrzött struktúráik miatt a purin SFC-k nagyon azonosak emberi társaikkal. Amellett, hogy ektodomainjeik 70%-ban azonosak, minden feltételezett glikozaminoglikán kötődési hely azonos. Ezenkívül a transzmembrán domén 96%-ban azonos, míg a citoplazmatikus domén 100%-ban azonos, beleértve három azonosan elhelyezkedő tirozinmaradékot [32]. Az emlős HSSPG-k szulfatációs mintázatának finom különbségei ellenére a konszenzusos HS-kötő motívumok, mint amilyenek a SARS-CoV-2 tüskéjében jelen vannak, lehetővé teszik a heparinkötő fehérjék emlősök HSPG-hez, köztük SFC-khez való kötődését [17,25,33]. Az SDC által közvetített endocitózis során a ligandumok kötődése kiváltja az SFC-k klaszterezését és serkenti az SDC–ligandum komplex internalizációját [20,34,35].
Tekintettel a SARS-CoV-2 HS-2 tüskék HS-kötő motívumának affinitására a HS-hez és a HSSPG-k megalapozott szerepére a nem beklasszikus endocitózisban, úgy döntöttünk, hogy elemezzük az inaktivált SARS-CoV-2 részecskék sejtfelvételét in vitro és in vivo egerek modelljeiben. A sejt- és állatkísérletek lehetővé tették a SARS-CoV-2 sejtbelépés fókuszált elemzését, és feltárták a SARS-CoV-2 internalizáció új aspektusait.
2. Eredmények
2.1. A SARS-CoV-2 internalizációjának elektronmikroszkópos vizsgálata purin sejtvonalakban
A hővel inaktivált SARS-CoV-2-t (törzs 2019-nCoV/USA-WA1/2020; lásd az 1. ábrátA,B ábra) két purin sejtvonallal – L929 fibroblasztokkal és RAW 264,7 (RAW) makrofágokkal – inkubálták 10, 30 és 180 percig 37 °C-on. A sejteket ezután rögzítettük, és a sejtek internalizálását elektronmikroszkóppal (EM) elemeztük, két rendszer felhasználásával: JEOL JEM-1400Flash és Delong LVEM 25

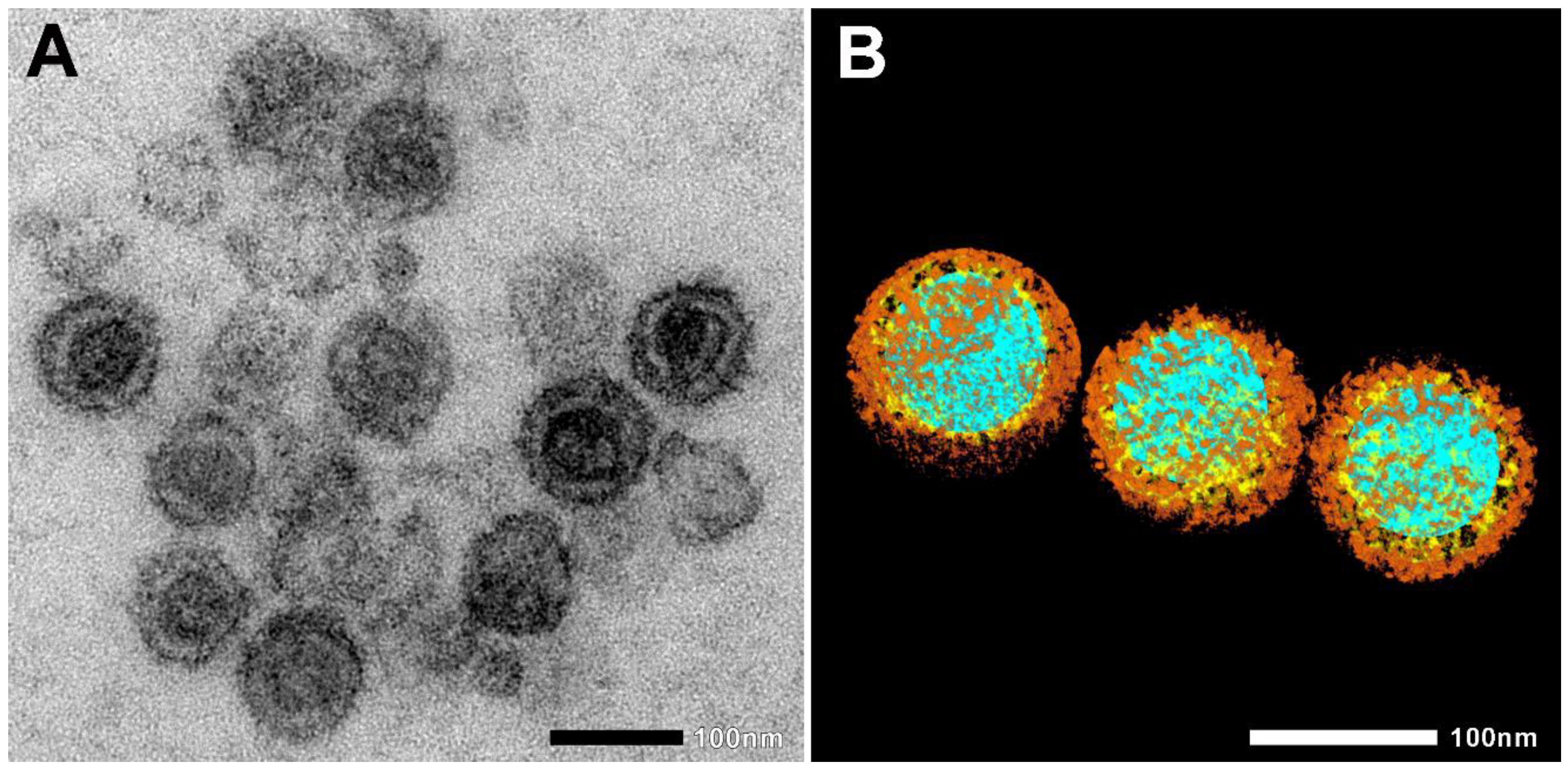
1. ábra. Elektronszkopikus képek hővel inaktivált SARS-CoV-2-ről. (A) Natív elektronmikroszkópos kép. (B) A SARS-CoV-2 morfológia megjelenítéséhez az érdeklődésre számot tartó régiókat, például a nukleinsavat (kék), a nukleokapszidot (sárga) és a tüskefehérjéket (barna) manuálisan szegmentáltuk. Skála sáv 100 nm.
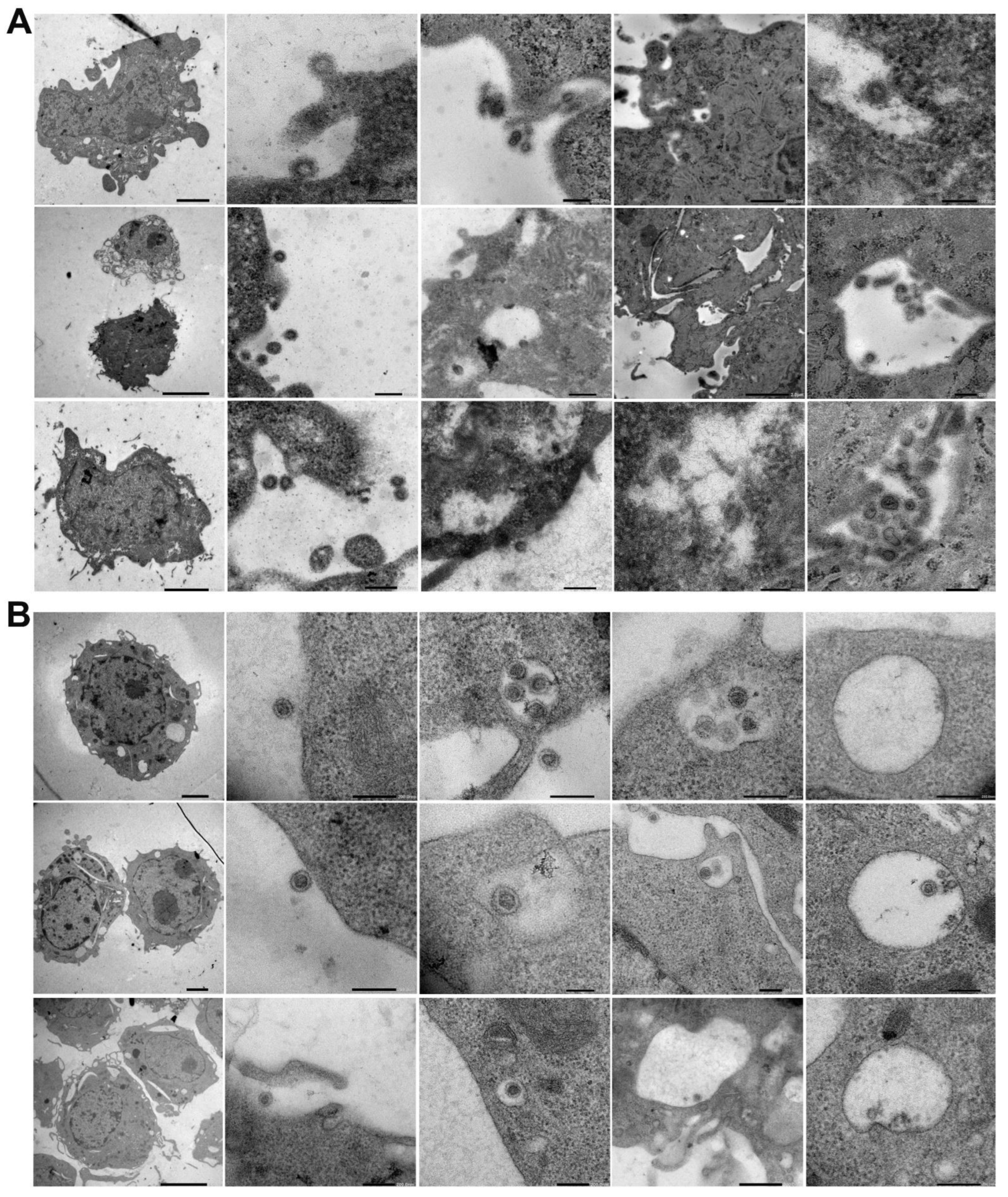
Az EM szépen feltárta a SARS-CoV-2 endocitózisának előrehaladását az L929 és RAW sejtekben a felszíni kötődéstől az endoszóma elnyeléséig (2. ábra, B). Így az EM kimutatta, hogy az ACE2 purin különbségei ellenére az inaktivált SARS-CoV-2 részecskéket a purinsejtek megköthetik és internalizálhatják (2. és 3. ábra, valamint S1 kiegészítő ábra).




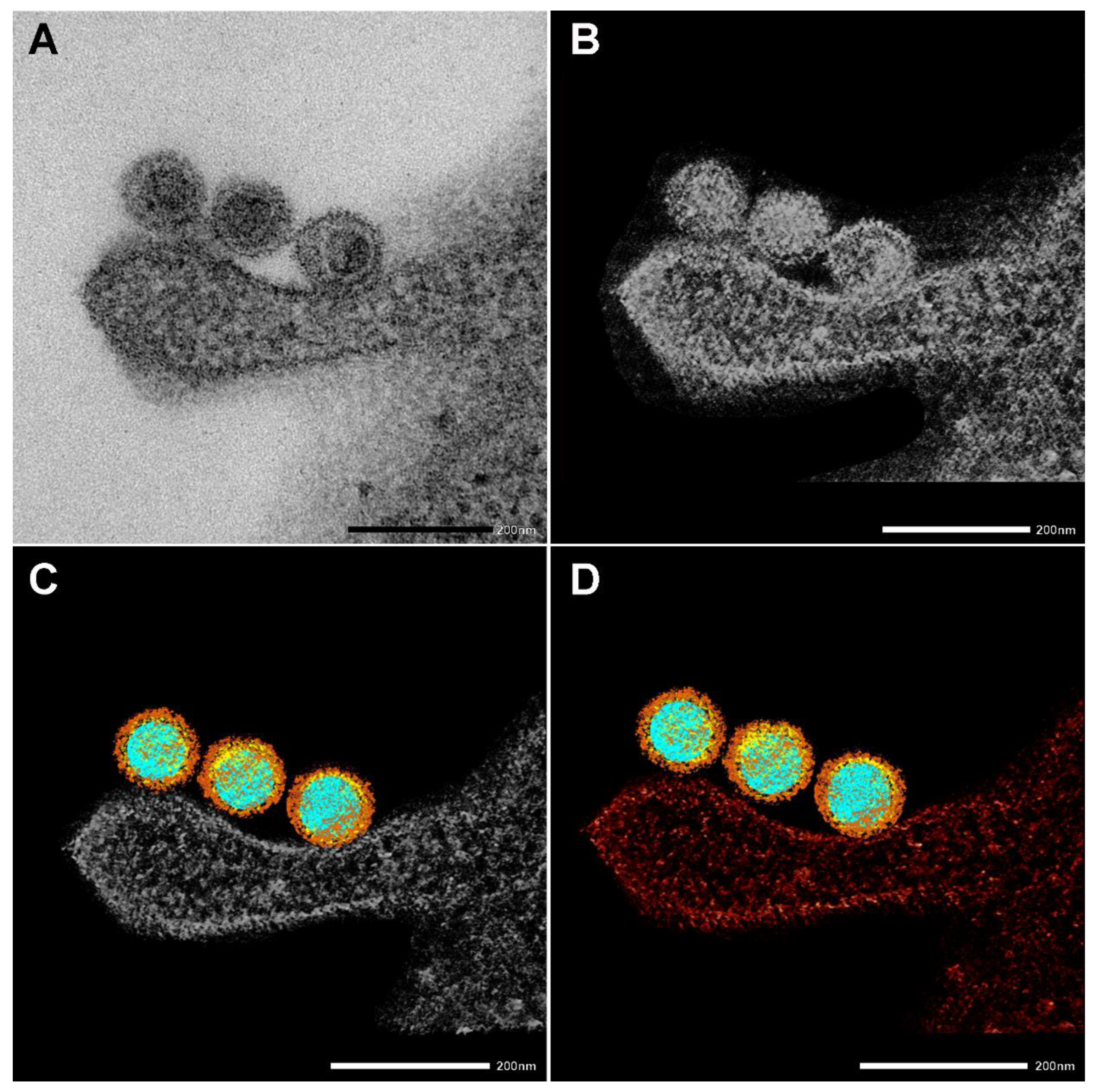
3. ábra. 3D elektron tomográfia létrehozása. (A) SARS-CoV-2 részecskék, amelyeket egy betolakodó RAW makrofogén köt össze 10 perc 37 °C-on történő inkubálás után. Skála sáv 200 nm. (B–D) A bemutatott képeket 3D-s elektron tomográfia létrehozására használják fel. Az érdeklődésre számot tartó régiókat, például a nukleinsavat (kék), a nukleokapszid (sárga) tüskefehérjéket (barna) és a szomszédos makrofágot (piros) manuálisan szegmentáltuk.
Az EM szépen ábrázolta a SARS-CoV-2 morfológiát és a vírus kölcsönhatását a sejtkomponensekkel. A 3. ábra a vírusoknak a betolakodó RAW makrofághoz való kötődésének elektron tomográfiájának létrehozásának négy szakaszát mutatja be (az így kapott 3D-s tomogramot a Grafikus absztrakt mutatja). Az egyes vírusrészecskék jellegzetes morfológiai jellemzőikkel megfigyelhetők, ahol a különböző színek képviselik a különböző régiókat. Az érdeklődésre számot tartó régiókat, például a nukleinsavat (kék), a nukleokapszidot (sárga) és a tüskefehérjéket (barna) manuálisan szegmentálták. Ezenkívül a szomszédos RAW makrofágot is szegmentálták és piros színnel ábrázolták.
2.2. A SARS-CoV-2 internalizációjának áramlási citometriai értékelése a purin sejtvonalakban
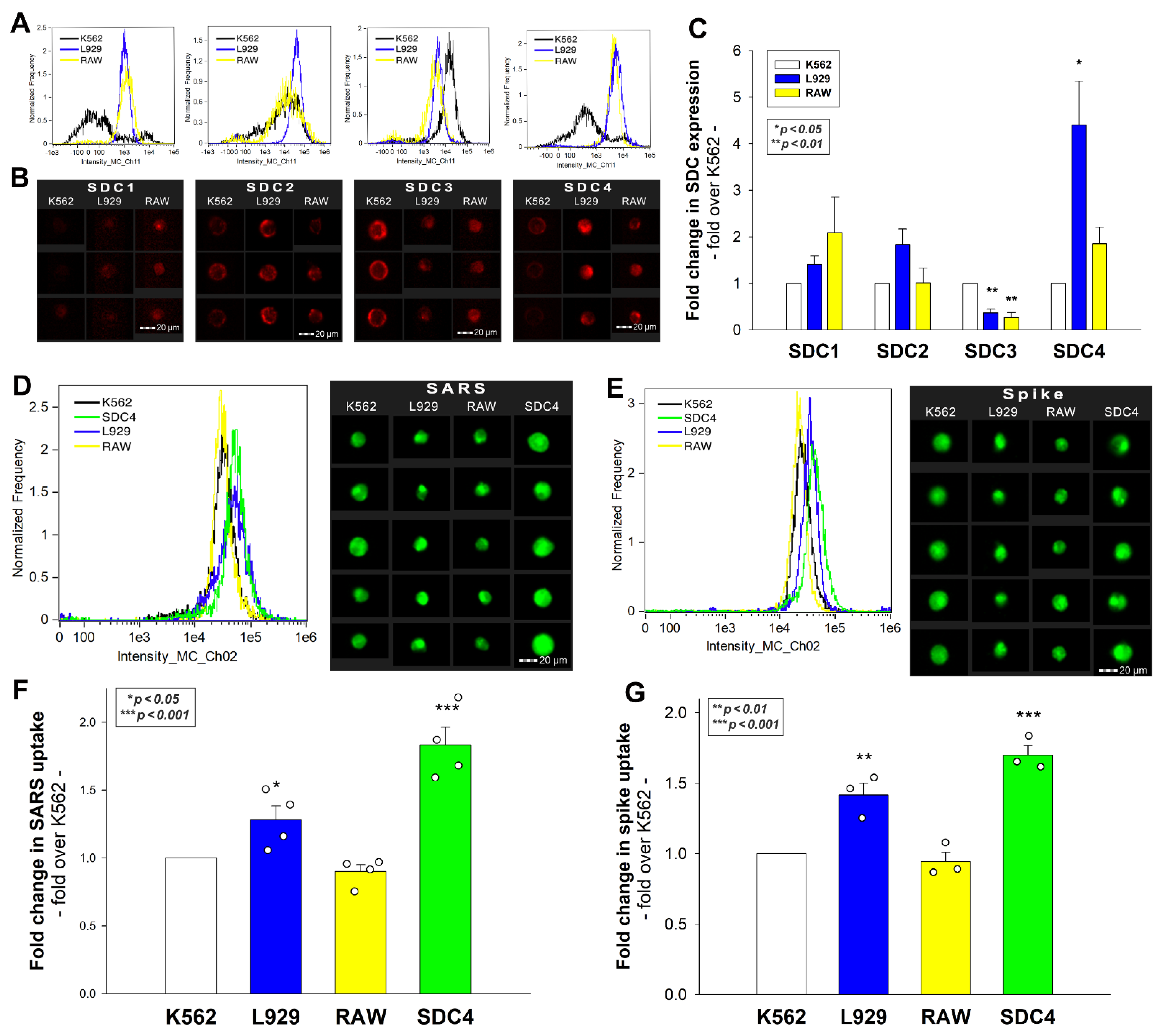
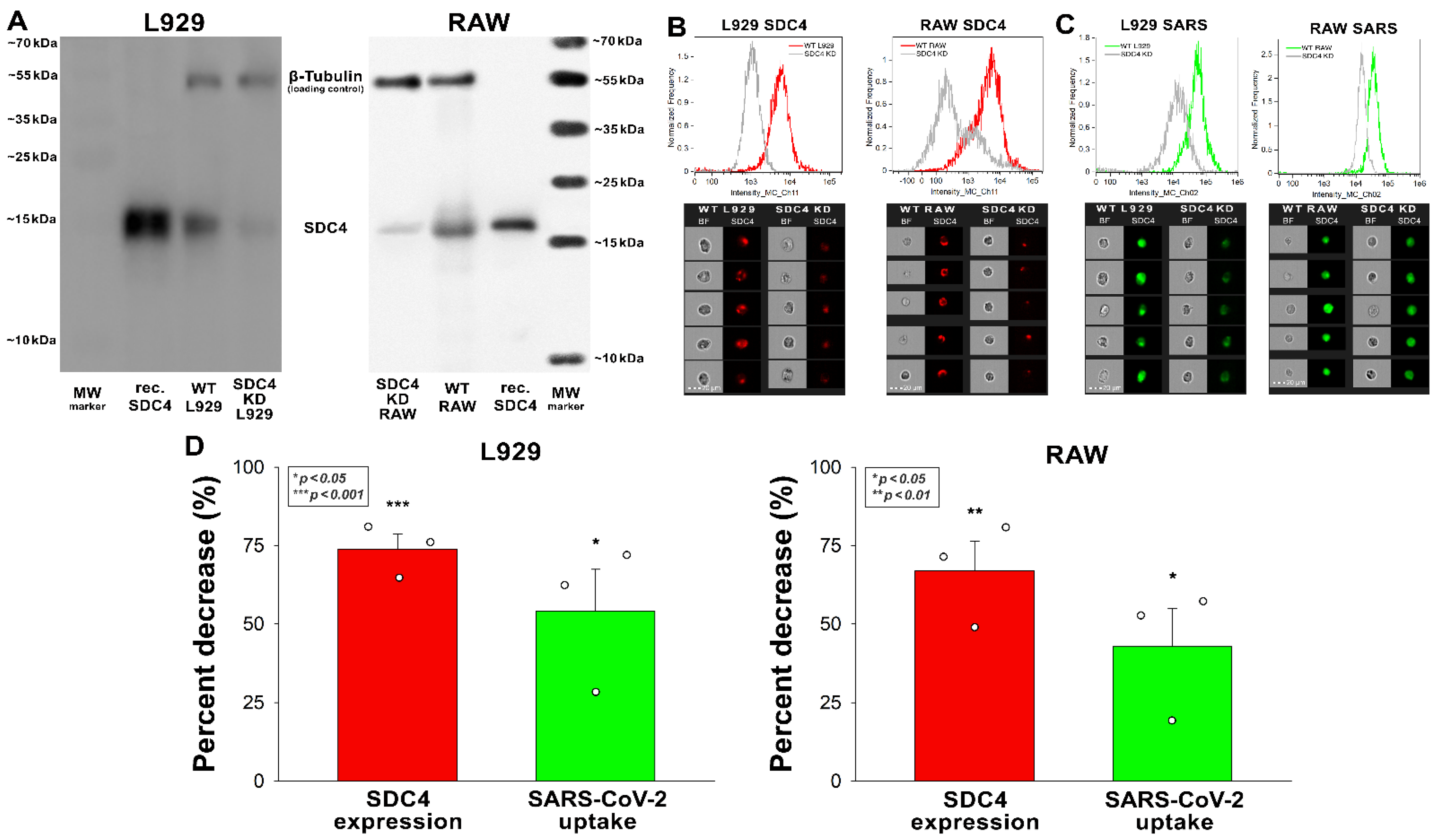
Korábban megmutattuk, hogy az SDC-k, különösen a tüdőben dúsított SDC4 közvetítik a SARS-CoV-2 sejt internalizációját az emberi sejtvonalakban. Miután feltártuk az inaktivált SARS-CoV-2 internalizálódását az EM-mel rendelkező purin sejtvonalakká, értékeltük az L929 és RAW sejtek SDC expresszióját. Mivel a K562 emberi sejtvonalból állítólag hiányoznak a HSMG-k, kivéve az endogén betaglikán és az SDC3 sekély mennyiségét, a K562 sejteket választották standardként [33,34,35]. A K562-höz képest az L929 és a RAW sejtek kissé megnövekedett SDC1-et mutattak, mégis jelentősen csökkent az SDC3 expressziója (3. ábra A–C). A K562-höz képest az L929 sejtek magasabb SDC2 expressziót mutattak, bár a növekedés statisztikailag nem volt szignifikáns (p > 0,05). A RAW és az L929 SDC4 expressziója szignifikánsan magasabb volt, mint a K562-é, de a növekedés (~4,5-szeres) határozottabb volt az L929 sejtekben. Az inaktivált SARS-CoV-2 sejtes internalizációja a RAW sejtekben valamivel alacsonyabb volt, mint a K562-ben, de az L929 fibroblasztok - amelyek SDC4 expressziója a legmagasabb volt a három vonal között - jelentősen megnövekedett SARS-CoV-2 felvételt mutattak (4.ábra D-F). A stabil SDC4 transzfektánsokhoz képest (amelyeket K562 sejtekben hoztak létre) ~20-szoros SDC4 expresszióval (S2 kiegészítő ábra), a SARS-CoV-2 sejtfelvétele az L929 sejtekben szignifikánsan alacsonyabb volt (4. ábraD-F). A tüskefehérje sejtes internalizálása (50 nM koncentrációban 3 órán keresztül 3 órán keresztül 37 °C-on inkubálva) a hővel inaktivált SARS-CoV-2 részecskékhez hasonló mintát mutatott (4.ábra E,G).


4. ábra. Az SDC expresszió és a SARS-CoV-2 áramlási citometriai értékelése, valamint tüskefehérje felvétele humán és purin sejtvonalakban. Az L929, RAW és K562 sejtek SDC expresszióját képalkotó áramlási citometriával elemeztük fluoreszcensen címkézett SDC antitestek felhasználásával. (A,B) Reprezentatív áramlási citometriai hisztogramok és fluoreszcens képek, amelyek az L929, RAW és K562 sejtek SDC expressziós szintjeit mutatják. Skála sáv = 20 μm. (C) Az észlelt SDC expressziós szintű cellákat K562-re normalizáltuk. A sávok három független kísérlet átlagát + SEM-jét képviselik. A statisztikai szignifikanciát a variancia (ANOVA) elemzésével értékeltük. * < 0,01 oldal; ** p < 0,001. (D–G) Az L929, RAW és K562 sejteket, valamint az SDC4 transzfektánsokat (amelyeket K562 sejtekben hoztak létre) inaktivált SARS-CoV-2-vel (1 MOI-nál) vagy tüskefehérjével (50 nM) inkubáltuk 3 órán keresztül 37 °C-on. Az inkubálás után a sejteket megmostuk, tripszinizáltuk, rögzítettük, permeabilizáltuk és kezeltük a tüske glikoproteinre (és az AF 488-címkével ellátott másodlagos antitestekre) specifikus antitestekkel. A SARS-CoV-2 és a tüske sejtfelvételét ezután képalkotó áramlási citometriával elemeztük. (D,E) Reprezentatív áramlási citometriai hisztogramok és fluoreszcens képek, amelyek a SARS-CoV-2- és tüskék által kezelt sejtek intracelluláris fluoreszcenciáját mutatják. Skála sáv = 20 μm. (F,G) Az észlelt fluoreszcencia intenzitásokat standardként SARS-CoV-2- vagy tüskekezelt K562 sejtekké normalizáltuk. A sávok négy (F) és három (G) független kísérlet átlag + SEM-jét képviselik. A statisztikai szignifikancia vs. szabványokat az ANOVA-val értékeltük. * p < 0,05; ** p < 0,01; p < 0,001.
A sejtek SARS-CoV-2-vel 4 °C-on történő inkubálása, a merev sejtmembránok miatt az endocitózist megszüntető hőmérséklet nagyon alacsony sejtfluoreszcenciát eredményezett (S3 kiegészítő ábra), így igazolva a SARS-CoV-2-vel kezelt sejteken 37 °C-on észlelt intracelluláris jelek megbízhatóságát. Érdemes megjegyezni, hogy a SARS-CoV-2 vagy a tüskefehérje-kezelés nem befolyásolta a sejtek életképességét (S4 kiegészítő ábra).
2.3. Az SDC4 leütés hatása a vírus internalizálására
Az L929 és RAW sejtek SDC4 expressziójának csökkentése leütéssel (KD) csökkentette a vírus sejtfelvételét (4 A–D ábra). Ez azt jelenti, hogy az SDC4 expresszió ~70%-os csökkenése ~50%-os csökkenést eredményezett a SARS-CoV-2 felvételben az alkalmazott egér sejtvonalakban (5.ábra B-D).

2.4. A SARS-CoV-2 internalizációjának konfokális mikroszkópos értékelése a purin sejtvonalakban
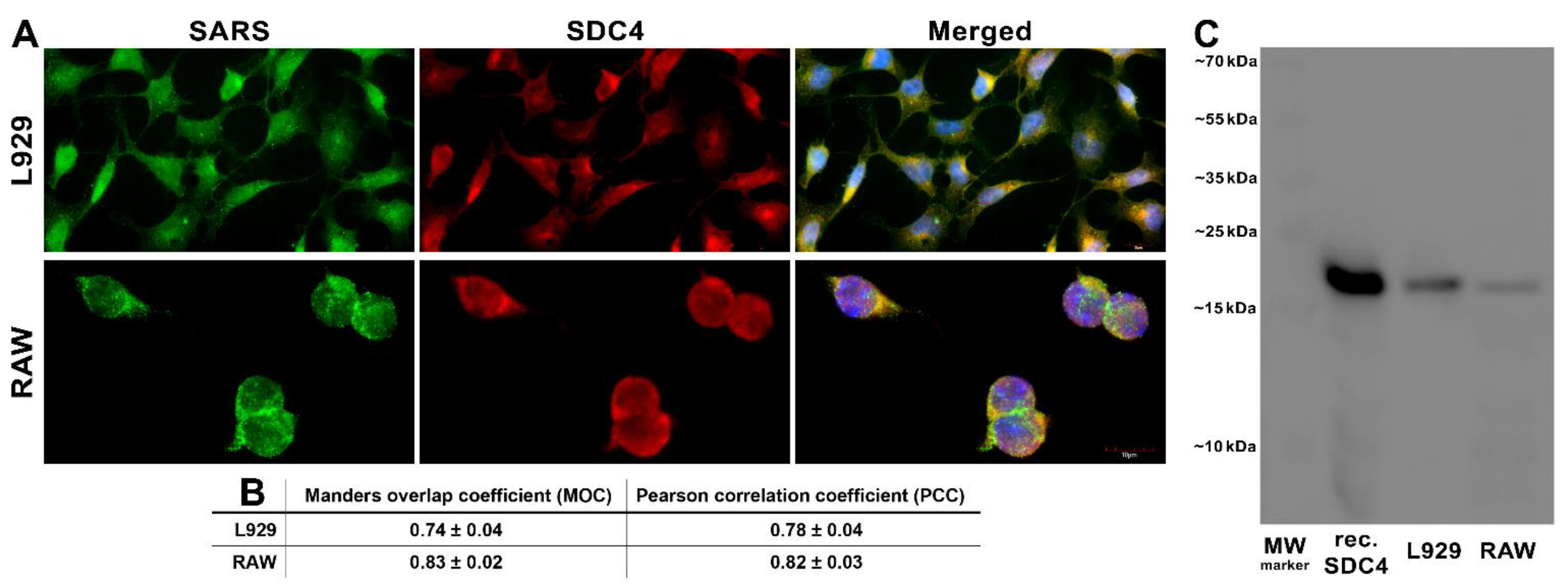
Ezután megvizsgáltuk a SARS-CoV-2 és az SDC4 kolokalizációját az L929 és RAW sejtekbe való felvétel során (6.ábra A, B ábra). Az inaktivált SARS-CoV-2-vel végzett 3 órás inkubálás során végzett konfokális mikroszkópos vizsgálatok nagy fokú átfedést mutattak ki (a 6. ábra A,B. ábrájána Manders átfedési együtthatójával (MOC) kifejezve) és kiváló korrelációt (lásd a 6B. ábraPearson-korrelációs együtthatóját (PCC)) a SARS-CoV-2 és az SDC4 között mindkét sejtvonalban (6 A,B ábra). A ko-immunprecipitáció azt is megerősítette, hogy az inaktivált SARS-CoV-2 kötődik az L929 és RAW sejtek SDC4-éhez (6C ábra).

6. ábra. A SARS-CoV-2 kollokalizálódik az SDC4-gyel a cellába való belépés során. Az L929 és RAW sejteket hővel inaktivált SARS-CoV-2-vel (1 MOI-nál) inkubáltuk 3 órán keresztül 37 °C-on. Az inkubálás után a sejteket mossuk, rögzítjük, permeabilizáljuk, és a tüske glikoproteinre (AF 488 címkézve) és az SDC4-re (APC címkével ellátott) specifikus antitestekkel kezeltük. A SARS-CoV-2 SDC-vel való kolokalizációját konfokális mikroszkóppal elemeztük. (A) A SARS-CoV-2 és az SDC kolokalizáció mikroszkópos elemzései. Három független kísérlet reprezentatív képei láthatók. Skála sáv = 10 μm. (B) A MOC és a PCC ± SEM-t az SDC és a SARS-CoV-2 átfedésére és kolokalizálására (a képek alatt jelezve) 18 kép elemzésével számítottuk ki, átlagosan 12 sejt elemzésével minden képen (3 különálló mintából). Skála sáv = 10 μm. (C) Reprezentatív nyugati folt, amely A SARS-CoV-2-vel immunprecipitált SDC4-et mutatja L929 és RAW sejtekben. 1. sáv: 0,5 μg SDC4; 2–3. sáv: SARS-CoV-2-vel kezelt L929 és RAW sejtek immunprecipititái. A standard fehérjeméret-markerek a jobb oldalon vannak feltüntetve.
Az inaktivált SARS-CoV-2-vel 4 °C-on inkubált sejtek – ez a hőmérséklet, amikor az endocitózis a sejtmembránok merevsége miatt leáll – viszonylag (azaz a 37 °C-on inkubált sejtekhez képest) alacsony felületalapú jeleket mutattak, a SARS-CoV-2 azonban még mindig jelentős átfedést (MCC) és korrelációt (PCC) mutatott az SDC4-gyel (S5 kiegészítő ábra). Az AF 488-as jelöléssel ellátott kecske anti-egér IgG - a SARS-CoV-2 kimutatására használt másodlagos antitest - és az APC-címkével ellátott SDC4 antitestek a vírus hiányában nagyon alacsony zöld autofluoreszcenciát mutattak (S6 kiegészítő ábra), így validálták a SARS-CoV-2-vel kezelt sejteken 37 ° C-on észlelt magas intracelluláris jeleket.
2.5. Inaktivált SARS-CoV-2 in vivo biodisztribuciója egerekben
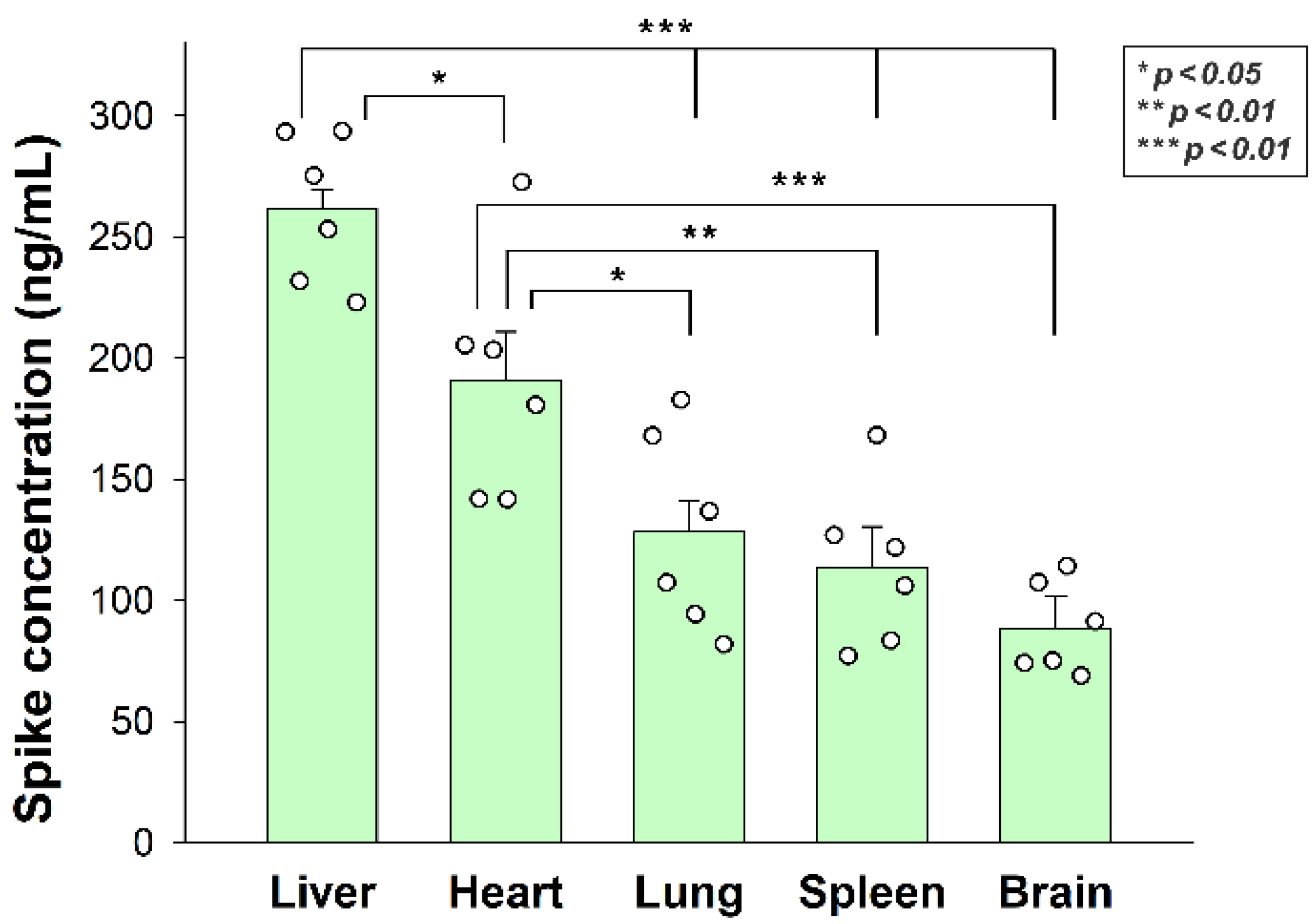
Az inaktivált SARS-CoV-2 in vitro sejtfelvételének értékelése után elemeztük annak in vivo biodisztribucióját WT egerekben. Az inaktivált SARS-CoV-2 részecskéket intravénásan (i.v.) fecskendeztük 12 hónapos C57BL/6 egerekbe. 3 órás inkubálás után az egereket érzéstelenítették, és a vért szívlyukasztással gyűjtötték össze. A jéghideg PBS-sel (2 ml/perc, 8 perc) végzett transzkardiális perfúzió után az agyat, a szívet, a májat, a tüdőt és a lépet - a Human Protein Atlas által SDC4 expressziójukra validált szerveket - eltávolították és szárazjégben fagyasztották le további vizsgálatok céljából [36,37,38]. A Human Protein Atlas szerint az SDC4 expresszió a májban, a tüdőben és az agyban a legmagasabb, míg a szívizom közepes, a lép pedig szerény SDC4 expressziót mutat (S1 kiegészítő táblázat). A SARS-CoV-2 internalizáció mértéke - a SARS-CoV-2 tüskespecifikus ELISA-val mérve - nemcsak azt az utat tükrözte, amelyet az i.v. beadott részecskék az adagolás után követnek, hanem az SDC4 szöveti expressziót is. Amint azt a 7. ábra mutatja, a máj lényegesen több SARS-CoV-2 részecskét vett fel, mint a többi szövet. A szív több SARS-CoV-2 részecskét is felhalmozódott, mint a tüdő, a lép és az agy. Az agy a vizsgált szervek közül a legkevésbé internalizálta a hővel inaktivált vírusrészecskéket. A tüdő és az agy közötti mért tüskekoncentrációk különbsége azonban nem volt szignifikáns, ami azt mutatja, hogy a SARS-CoV-2 jelentős mennyisége jut be az agyba (7. ábra). A kezeletlen kontrollok szerveiben a tüskekoncentrációk marginálisak voltak, és körülbelül a SARS-CoV-2-vel kezelt állatok eszközeinek standard hibájának tartományába tartoztak (S7 kiegészítő ábra).

7. ábra. Az inaktivált SARS-CoV-2 in vivo internalizálása egerekben. A C57BL/6 egereket hővel inaktivált SARS-CoV-2-vel kezeltük. 3 órás inkubálás után az egereket eutanizáltuk, különböző szerveket gyűjtöttünk össze, és SARS-CoV-2 tüskekoncentrációjukat tüskespecifikus ELISA készlettel mértük. Minden csoport 6 állatot tartalmazott. A sávok az átlag + SEM-t képviselik. * p < 0,05; ** p < 0,01; p < 0,001.
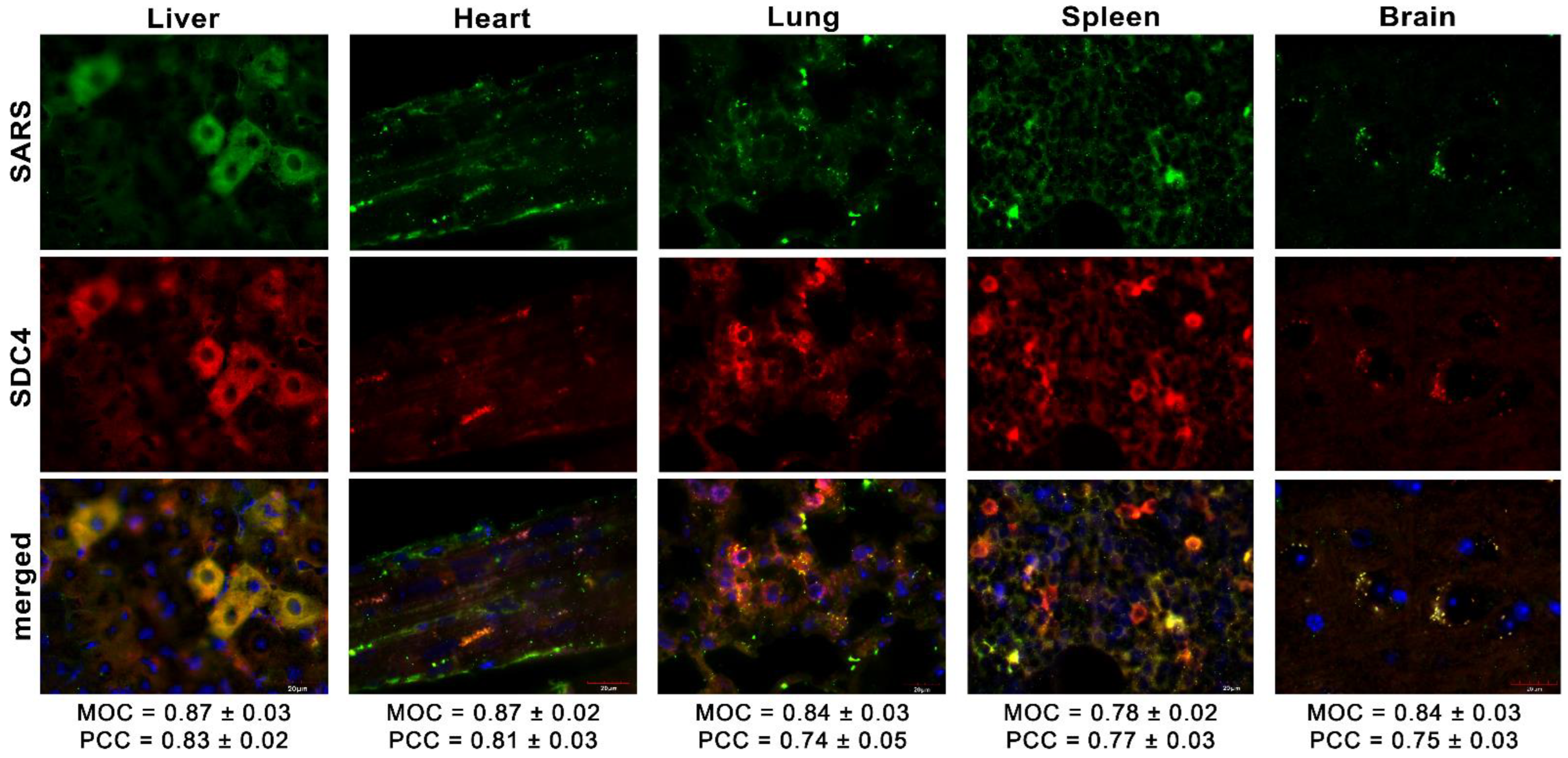
Ezzel párhuzamosan megvizsgáltuk az inaktivált SARS-CoV-2 in vivo internalizálását konfokális mikroszkópiával. Ezek a mikroszkópos elemzések nagy átfedést mutattak (lásd a 8. ábrán látható MOC-okat), valamint kiváló korrelációt mutattak az internalizált SARS-CoV-2 és az SDC4 között. A SARS-CoV-2-vel kezelt állatokkal ellentétben a hisztokémia nem tárta fel a SARS-CoV-2 jelenlétét a SARS-CoV-2 kezelésben nem részesülő kontrollállatokban (S8 kiegészítő ábra).

8. ábra. Az SDC4 és a SARS-CoV-2 kolokalizációja az egerek különböző szerveiben. Máj-, szív-, tüdő-, lép- és agyi minták olyan egerekből, amelyek i.v. inaktivált SARS-CoV-2-t kapnak. A szövetI SDC4-et APC-címkével ellátott SDC4 antitesttel, míg a SARS-CoV-2-t AF 488-as tüske antitesttel detektáljuk. Skála sáv = 20 μm. A MOC ± SEM és a PCC ± SEM a SARS-CoV-2 és az SDC4 átfedésére és kolokalizációjára vonatkozó MOC-t úgy számítottuk ki, hogy minden állat 3 mintájából 18 képet elemeztünk.
2.6. Az inaktivált SARS-CoV-2 megemeli a szövet tnf-α koncentrációját
Amint azt a TNF-α-specifikus ELISA-analízisek feltárták, az internalizált SARS-CoV-2 részecskék jelentősen megemelték a szöveti TNF-α koncentrációját minden szervben (agy, szív, máj, tüdő és lép). Amint azt a 9. ábra mutatja, az inaktivált vírusrészecskék váltották ki a szív legmagasabb TNF-α koncentrációját, amelyet a tüdő, a lép, a máj és az agy követett. Így az inaktivált SARS-CoV-2 proinflammatorikus útvonalakat vált ki minden szervben, amelybe belép. A SARS-CoV-2 által kiváltott szöveti gyulladás az adott szövet által felhalmozódott vírusrészecskék számához kapcsolódik, kivéve a májat, amely a legjelentősebb számú vírusrészecskét gyűjtötte össze. A máj TNF-α szintje azonban hasonló volt az agyéhoz.

3. Összefoglalás
Több mint két és fél évvel a SARS-CoV-2 megjelenése után még mindig sok a megválaszolatlan kérdés a SARS-CoV-2 sejtbiológiája körül [39,40]. A COVID-19 patomechanizmus egyik legjelentősebb kihívása annak megértése, hogy a SARS-CoV-2 sejtes belépése hogyan járul hozzá a fertőzéshez [41]. Ehhez meg kell kérdőjelezni az ACE2 kizárólagosságát a SARS-CoV-2 sejtes internalizálásában. Ez különösen igaz, ha a tudományos bizonyítékok egyre nagyobb száma alátámasztja a SARS-CoV-2 ACE2-független sejtes bejegyzését [7,8,9]. Megvizsgáltuk a SARS-CoV-2 SDC-függő bejegyzését és Delta változatát. Tanulmányaink szerint az SDC-k - különösen az SDC4, a tüdőben dúsított izoforma - megkönnyítik a SARS-CoV-2 és a Delta variáns internalizációját tüskefehérjéik rögzítésével [18,24]. A csökkenő SDC expresszió jelentősebben csökkentette a SARS-CoV-2 sejtfelvételét, mint az ACE2 [18,24]. Korábbi vizsgálataink azt is kimutatták, hogy az ACE2 aktivitása nem szükséges a vírus hatékony sejttranzokációjához [18,24]. Ez azt jelenti, hogy az ACE2 gátlása kisebb hatással volt a SARS-CoV-2 sejtfelvételére, és egyik sem a Delta variánsára, ami arra utal, hogy az ACE2 dokkolóhelyként szolgál, nem pedig endocitózis receptorként, amely a membrán elnyelését és a vírus internalizációját hajtja [24]. Az SFC-k mellett más sejtes SARS-CoV-2 receptorokat is azonosítottak, amelyek lehetővé teszik a vírus ACE2-független sejtfelvételét [7,8,9].
Az emberi és a purin ACE2 közötti különbségek miatt a WT egerek, mint a Muridae család más rágcsálófajai, állítólag nem fertőzhetők MEG SARS-CoV-2-vel [42]. Az egerek azonban még mindig HSMG-ket fejeznek ki, beleértve az SFC-ket is, lehetővé téve bizonyos ligandumok, köztük a vírusok és más paraziták kötődését és hatékony internalizálását [43,44]. A SARS-CoV-2 tüskéje nagy affinitással bír a HS iránt a konszenzusos heparin-kötő motívum (PRRAR) miatt [13,16,18,24]. Ugyanez a motívum lehetővé teszi az emberi fibronektin kötődését patkány HSSPG-khez, beleértve az SFC-ket és a glicánokat is, a patkány és az emberi HSPG-k közötti kisebb strukturális különbségek ellenére [17].
Figyelembe véve az evolúciósan megőrzött heparinkötő motívum jelenlétét a SARS-CoV-2 tüskéjében, valamint az emlős HSMG-k részvételét a ligandumok sejtfelvételében, in vitro és in vivo vizsgálatokat végeztünk az inaktivált SARS-CoV-2 részecskék internalizálásáról. Eredményeink azt mutatják, hogy az inaktivált SARS-CoV-2 részecskék hatékonyan internalizálódnak a purin sejtvonalakba, megalapozott SDC4 expresszióval. Az inaktivált SARS-CoV-2 részecskék kollokalizálódnak az SDC4-gyel a sejtek felvétele során, ami közös belépési útvonalra utal. Az i.v. in vivo beadása után a vírusrészecskéket az egerek különböző szervei veszik fel, beleértve a májat, a szívet, a tüdőt, a lépet és az agyat. Az internalizált SARS-CoV-2 részecskék kiváló átfedést mutatnak az SDC4 szövetekkel, megerősítve az SDC4 szerepét a vírus in vivo sejtfelvételének közvetítésében. Érdemes hangsúlyozni a vírusrészecskék megjelenését az agyban is. Az SFC-kről ismert, hogy megkönnyítik a HIV-1 behatolását az agyba [45]. Így még a strukturálisan inkompetens ACE2 esetében is, a meglévő sejtes belépési útvonalak hatékony SARS-CoV-2 szállítást kínálnak az agyba. A szervekbe való belépés után az internalizált SARS-CoV-2 részecskék növelik a szöveti TNF-α koncentrációkat, ami helyi gyulladás kialakulására utal. Így az immunmechanizmusok könnyen kimutatják az idegen vírusrészecskéket a WT egerekben.
Így adataink megerősítik a SARS-CoV-2 SDC4 által közvetített sejtbejára vonatkozó korábbi eredményeket, ami arra utal, hogy még egy strukturálisan eltérő ACE2 murine esetében is az SDC4 - egy jól megalapozott endocitikus funkciókkal rendelkező receptor - még mindig képes megkönnyíteni a vírus sejtfelvételét. Figyelembe véve, hogy ugyanaz a heparinkötő motívum, amely a SARS-CoV-2 tüskéjében jelen van, fibronektint ad a rágcsáló SFC-khez való kötődéshez, a SARS-CoV-2 sejtes belépése az SFC-ket kifejező purinsejtekbe, különösen az SDC4-be, nem meglepő. A SARS-CoV-2 számos endocitásos mechanizmusa ki volt téve [46,47]. Ezek közül a klatrin által közvetített endocitózist javasolták a vírus fertőzőképességének kulcsfontosságú aspektusának [48]. A SARS-CoV-2, csakúgy, mint más β koronavírusok, lizoszómákat használ a kijutáshoz, ezáltal bizonyítva az endoszomális /lizoszomális rendszer fontosságát a SARS-CoV-2 életciklusa és fertőzése szempontjából [48,49,50,51]. Az SDC által közvetített sejtbejárat elsősorban klatrinfüggetlen útvonalakon keresztül történik, lipidtutajokkal [52]. A klatrintól független SDC által közvetített útvonalakon keresztül a sejtekbe belépő ligandumok megkerülhetik a lizoszomális lebomlást [52,53,54,55,56]. Így a lizoszómák elkerülésével a SARS-CoV-2 SDC által közvetített internalizálása paradox módon gátolhatja a lizoszómákból kilépő új vírusok kialakulását. Mivel a β-koronavírusok eltérítik az endolizomoszóma-rendszert a fertőzés biztosítása érdekében, a lizoszomális belépést elkerülő endocytikus útvonalak kevésbé kedvező miliőt kínálnak a SARS-CoV-2 replikációhoz. A vírusciklus hat fázisból áll: (1) gazdasejt-kötődés, (2) gazdasejt-belépés, (3) bevonat nélküli, (4) replikáció, (5) összeszerelés és (6) felszabadulás a sejtből [51]. Még akkor is, ha a sejtek kötődése és belépése hatékony, a hiányos vírusreplikáció, összeszerelés és felszabadulás alááshatja a fertőzés hatékonyságát. Mivel az SDC által közvetített endocitózis, a klasszikus klatrin által közvetített útvonalakkal ellentétben, inkább elkerüli a lizoszomális belépést, az inaktivált vírus hatékony sejtfelvétele nem mond ellent a hatékony SARS-CoV-2 fertőzés hiányának wt egerekben. Másrészt, még a nem hatékony replikáció esetén is, a vírusrészecskék még mindig gyulladásos útvonalakat válthatnak ki WT egerekben, ami egyértelmű jele az idegen vírusrészecskék immunfelismerésének.
Összefoglalva, adataink azt mutatják, hogy a SARS-CoV-2 részecskék hatékonyan bejutnak wt egerekbe, és megerősítik a SARS-CoV-2 ACE2-független belépési útvonalaira vonatkozó, nemrégiben feltárt bizonyítékokat. A kapott bizonyítékok alátámasztják az endocita útvonalaknak a vírusfertőzésre és a replikációra gyakorolt nagy hatását is. Így jelenlegi kéziratunk számos kritikus megállapítást tartalmaz a SARS-CoV-2-ről, hogy jobban megértsük a vírus sejtbiológiáját.
4. Anyagok és módszerek
4.1. Hővel inaktivált SARS-CoV-2
A hővel inaktivált SARS-CoV-2-t (törzs: 2019-nCoV/USA-WA1/2020) az ATCC-től (Manassas, VA, USA; cat. no. ATCC VR-1986HK).
4.2. SDC4 konstrukciók és transzfekció, sejttenyészetek
SDC4 transzfektánsok, K562 sejtekben (ATCC, Manassas, VA, USA; cat. no. CCL-243), a korábban leírtak szerint jöttek létre [18]. A stabil SDC4 transzfektánsokat úgy választottuk ki, hogy az SDC4 expressziót áramlási citometriával mértük APC-jelöléssel ellátott SDC4 antitestek felhasználásával (R&D Systems, Minneapolis, MN, USA; SDC4: monoklonális patkány IgG2a klón #336304, macska. Nem. FAB29181A [20,54]), a megfelelő izotípus-szabályozással együtt (K+D rendszerek; patkány IgG2a APC izotípus-szabályozás, macskaszám. IC006A).
L929 (Merck KGaA, Darmstadt, Németország; macskaszám: 85011425-1VL) és RAW 264,7 sejtek (ATCC, Manassas, VA, USA, cat. no. TIB-71) DMEM-ben termesztették, 10% FBS 37 °C-on, 5% CO2.
4.3. SARS-CoV-2 sejtes kötődés és internalizálás elektronmikroszkópos vizsgálata
Az L929 és RAW sejteket üvegfenéki 35 mm-es tenyészedényeken termesztettük. 24 óra elteltével a sejteket SARS-CoV-2-vel (1 MOI) inkubáltuk különböző ideig (10, 30 és 180 perc) DMEM /F12 táptalajban (10% FBS-szel kiegészítve) 37 °C-on. A SARS-CoV-2-vel vagy anélküli inkubálás után a sejteket kétszer öblítettük jéghideg PBS-sel, rögzítve egy Karnovszkij-oldatban, amely 2% paraformaldehidet (Sigma; St. Louis, MO, USA) és 2,5% glutaraldehidet (Polysciences; Warrington, PA, USA) foszfátpufferben egy éjszakán át 4 °C-on [57,58]. Ezután a mintákat desztillált vízben (pH 7,4) öblítettük le, és 60 perc osmifikáció után 2% OsO-val4 oldatban (desztillált vízben, pH 7,4) és 10 percig desztillált vízben történő ismételt öblítéssel a mintákat osztályozott etanolsorozaton keresztül dehidratáltuk (50%–100%, minden koncentrációban 10 percig), és propilén-oxidon [58,59] keresztül folytattuk. A dehidratáció után a mintákat egy epoxi alapú műanyagba ágyaztuk be (Durcupan ACM; Sigma), és polimerizálva 56 °C-on 48 órán keresztül [58]. A műanyag tömbökből 50 nm ultravékony metszetet vágtunk ki ultracut UCT ultramicrotome-val (Leica; Wetzlar, Németország) vagy egy RMC Boeckler PTPCZ Ultramicrotome (RMC Boeckler, Boeckeler Instruments, Inc., Tucson, AZ, USA). Az ultravékony metszeteket ezután egylyukú, formvarral bevont rézrácsokra helyezték (Electron Microscopy Sciences; Hatfield, PA, USA), ellentétben a 2% uranil-acetáttal (Elektronmikroszkópiás Tudományok) 50% etanolban (Molar) és 2% ólom-citrátban (Elektronmikroszkópos Tudományok) desztillált vízben (Reynolds, 1963; Hayat, 1970). A mintákat ezután két külön mikroszkópos laboratóriummal átvizsgálták két transzmissziós elektronmikroszkóppal: egy JEM-1400Flash -rel (JEOL; Tokió, Japán) és egy Delong LVEM 25 (Delong Instruments a.s., Brno, Csehország).
4.4. Elektronmikroszkópos tomográfia
Az elektron tomográfiához a korábban leírt 150 négyzet alakú mash rácson elkészített mintákat egy nagy dőlésszögű EM tartófejbe (JEOL; Tokió, Japán); majd kiválasztották a tomográfiára alkalmas területeket. Minden szögből egyetlen képet rögzítettünk a Recorder for TEM (System in Frontier; Tokió, Japán) −44 és +44 billentési szög között. A kézi fókuszt és az automatikus pozíciókövetést a teljes dőléssorozattal együtt használták. A képkötegfájlokat tmg fájlformátumba mentettük a további rekonstrukcióhoz, ahol a képeket egy igazított képblokkba építették be a Composer (System in Frontier) segítségével. Miután kiválasztották az előválasztás érdeklődési területét, a fiduciális ultrastruktúrát, például az éleket és a nagy kontrasztkülönbséggel rendelkező területeket jelölték meg; majd keresztkorrelációs igazítást és sinogram alapú szűrt visszavetítést végeztünk. Az inverz fekete-fehér bin fájlként exportált rekonstruált blokkot a Visualizer-Kai -ban (System in Frontier) dolgoztuk fel. A min,max,gamma és alfa értékeket színátmenet módban állítottuk be a rekonstruált átlátszatlan térkép legjobb megjelenítése érdekében. Az összes vizualizált régiót (sejtterület, teljes részecsketerületek, tüskefehérjék és nukleokapszid) manuálisan szegmentálták a Visualizer-Kai ROI alösszetevőjében, új kötetekbe mentették, és egyszínű térképekkel színezték a jobb megjelenítés érdekében.
4.5. Az SDC expresszió áramlási citometriás analízise
Az alkalmazott sejtvonalak (K562 sejtek, SDC4 transzfektánsok, L929 és RAW 264,7) SDC expresszióját áramlási citometriával mértük APC-jelöléssel ellátott anti-humán SDC4 (R&D Systems, Minneapolis, MN, USA; SDC4: monoklonális patkány IgG2a klón #336304, macska. Nem. FAB29181A) vagy primer anti-murine SDC antitestek (egér SDC1, cat. no. AF3190, K+F rendszerek, Minneapolis, MN, USA; egér SDC2, macska. Nem. PA5-95938, Invitrogen, Waltham, MA, USA; egér SDC3, macska. Nem. PA5-47377, Invitrogen, Waltham, MA, USA; egér SDC4-APC címkézett antitest, macska. no. 130-109-831, Miltenyi Biotec, Bergisch Gladbach, Németország) és megfelelő másodlagos antitestek (nyúl anti-kecske IgG (H+L) kereszt-adszorbeált másodlagos antitest, Alexa Fluor™ 633, cat. no. A-21086, Invitrogen, Waltham, MA, USA; kecske anti-nyúl IgG (H +L) kereszt-adszorbeált másodlagos antitest, Alexa Fluor™ 633, macska. Nem. A-21070, Invitrogen, Waltham, MA, USA) és a megfelelő izotípus-vezérlők a gyártó protokollja szerint.
4.6. SARS-CoV-2 felvétel áramlási citometriai analízise
A WT K562, L929, RAW 264.7 és SDC4 transzfektánsokat az inaktivált SARS-CoV-2 internalizálásának számszerűsítésére használták. Röviden, 6 × 105 DMEM/F12 táptalajban lévő sejteket/ml-t SARS-CoV-2-vel (1 MOI-nál) inkubáltuk 3 órán keresztül 3 órán keresztül 37 °C-on. 3 órás inkubálás után a sejteket tripszinizáltuk (a Nakase et al. [60,61]) által leírt módszerrel), hogy eltávolítsuk az extracellulárisan kapcsolódó vírusrészecskéket a sejtfelszínről. Ezután a sejteket szobahőmérsékleten 1 órán át mossuk, rögzítjük, permeabilizáljuk és a megfelelő szérummal blokkoltuk. A sejteket ezután egér monoklonális (1A9) és SARS tüskés glikoprotein (Abcam, Cambridge, UK, cat. no. 273433) kezelték, majd AF 488-as címkével ellátott kecske anti-egér IgG-vel (Invitrogen, Carlsbad, CA, USA, cat. no. A-11001). A mintákat ezután háromszor leöblítettük 1% BSA-t és 0,1% Triton X-100-at tartalmazó PBS-sel, és az áramlási citometria felé haladtunk. A sejtek felvételét ezután áramlási citometriával mértük egy Amnis FlowSight képalkotó áramlási citométerrel (Amnis Corporation, Seattle, WA, USA). Mintánként legalább 5000 eseményt elemeztünk. Az előre-szóró-oldal-szórás ábrázolás megfelelő kapuzását alkalmazták a sejttörmelék és az aggregátumok kizárására. A fluoreszcencia analízist az Amnis IDEAS elemző szoftverrel végeztük.
4.7. SARS-CoV-2 felvétel áramlási citometriai analízise
Az SDC4 leütést (KD) L929 és RAW sejtekben az egér SDC4 shRNS-re specifikus lentivirális vektorrendszerrel (cat. no. sc-36589-SH) a gyártó protokollja szerint (Santa Cruz Biotechnology, Inc., Dallas, TX, USA). A stabil KD sejteket 2 mg G418-ban választottuk ki, és képalkotó áramlási citometriával (Amnis FlowSight, Luminex Corporation, Austin, TX, USA) válogattuk össze anti-SDC4 antitesttel és megfelelő izotípus-szabályozással, a fent leírtak szerint. Az SDC4 sejtes expresszióját a leütést követően a nyugati blottinggal is meghatároztuk, amint azt korábban leírtuk [62].
4.8. Az in vitro kolokalizáció konfokális mikroszkópiája
A SARS-CoV-2 és az SDC4 együttes elhelyezését L929-ben és RAW 264.7 sejtekben konfokális mikroszkóppal jeleztük. Az L929 és RAW 264,7 sejteket poli-D-lizinnel bevont üvegfedezetű, 35 mm-es tenyészedényeken (MatTek Corp., Ashland, MA, USA) termesztettük. 24 óra elteltével a sejteket SARS-CoV-2-vel (1 MOI) inkubáltuk 3 órán keresztül 37 °C-on. A sejteket ezután kétszer öblítettük jéghideg PBS-sel, és 4% -os paraformaldehidben rögzítettük (Sigma, St. Louis, MO, USA), a sejtmembránokat permeabilizáltuk (0,1% Triton X-100), és a sejteket szobahőmérsékleten 1 órán keresztül a megfelelő szérummal blokkoltuk, majd az áramlási citometriai elemzésekhez leírt specifikus S1 és SDC4 antitestkezeléseket. A mintákat ezután háromszor leöblítettük 0,1% Triton X-100-at tartalmazó PBS-sel, majd DAPI-vel (1:5000) 15 percig megfestettük, HÁROMSZOR megmostuk PBS-sel, háromszor megmostuk PBS-sel, és beágyaztuk a Fluoromount G-be (SouthernBiotech, Birmingham, AL, USA) [20,54]. A fluoreszcens eloszlást ezután egy Aurox Clarity lézermentes konfokális egységgel felszerelt Leica DMi8 mikroszkóppal elemeztük. A bemutatott metszeteket körülbelül a sejtek éjféli szintjén vettük. A fotomultiplier nyereség és a megvilágítási teljesítmény minden kísérleten belül azonos volt. Az Aurox Visionary szoftvert konfokális mikroszkóppal történő képszerzésre használták. A kolokalizációs elemzésekhez a képeket az ImageJ (NIH, Bethesda, MD, USA) JACoP beépülő moduljával elemeztük.
4.9. A sejtek életképességének mérése
A SARS-CoV-2 és a tüske sejtéletképességre gyakorolt hatását az EZ4U sejtproliferációs vizsgálattal értékeltük (Biomedica GmBH, Bécs, Ausztria, cat. no. BI-5000) a gyártó utasításai szerint. Az abszorbanciát BioTek Cytation 3 multimódusú mikrolemez olvasóval mértük.
4.10. Állatkísérletek
2 × 106 inaktivált SARS-CoV-2 részecskék 200 ml PBS-ben (Lonza, Bázel, Svájc; cat. no. BE17-516F) intravénásan (azaz v.) adták be 12 hónapos C57BL/6 egerekbe. A kontroll egerek 200 ml tiszta PBS (azaz vírust nem tartalmazó) injekciót kaptak, azaz nem kaptak injekciót. Minden csoport (azaz a kezelt SARS-CoV-2 és a kontrollok) 6 állatból állt. 3 órás inkubálás után az egereket 2,2,2-tribróm-etanollal (cat. no. T48402, Merck KGaA, Darmstadt, Németország), és a vért szívdúcióval gyűjtötték össze. Jéghideg PBS-sel (2 ml/perc, 8 perc) végzett transzkardiális perfúzió után az agyat, a szívet, a májat, a tüdőt és a lépet eltávolították és szárazjégben fagyasztották további vizsgálatok céljából.
4.11. A SARS-CoV-2 tüske egérszöveti koncentrációjának mérése
Az agy-, szív-, tüdő-, máj- és lépmintákat homogenizáltuk lízis pufferben (Qiagen) 1% NP-40/PBS-ben egy Complete Mini EDTA-mentes proteázgátló koktélban (Roche). A szöveti lizátumokat RayBio COVID-19/SARS-CoV-2 Spike Protein ELISA készlettel elemeztük (cat. no. ELV-COVID19S2, RayBiotech, Inc., Peachtree Corners, GA, USA) a gyártó utasításai szerint.®
4.12. Az egérszövetminták immunhisztokémiája
Immunhisztokémia esetén egérszöveti mintákat (n = 6 egér csoportonként) 18 órán át rögzítettünk 4% paraformaldehidben (cat. no. P6148, Sigma-Aldrich), majd etanolsorozatban dehidratálva, xilollal (kb. 00699464, Avantor Inc., Radnor, PA, USA) tisztítva és paraffinba ágyazva (26154.291, Avantor Inc.). A 10 μm vastag metszet mennyiségét végül mikrotómával vágtuk (Leica Biosystems Inc., Buffalo Grove, IL, USA), és a szakaszokat SuperFrost Plus diákon gyűjtötték össze (Thermo Fisher Scientific Inc., Waltham, MA, USA). Az antigén kimutatását hő okozta antigén visszanyeréssel végeztük. A diákat először 10 percig 95–100 fokra melegített citrátpufferbe merítettük, majd kb. 20 percig szobahőmérsékletre hűtöttük. Ezután a tárgylemezeket blokkoló oldatba helyeztük (0,1% PBST-vel hígított 5% -os kecske- vagy szamárszérum) szobahőmérsékleten 30 percig, majd a blokkoló oldatot öblítés nélkül eltávolítottuk. A tárgylemezeket ezután primer antitestekkel (egér monoklonális, 1A9, SARS tüske glikoprotein, Abcam, Cambridge, Egyesült Királyság, 8. kat 273433.) és APC-jelöléssel ellátott purin-ellenes SDC4 antitestekkel (macskaszám: 130-109-831, Miltenyi Biotec, Bergisch Gladbach, Németország) inkubáltuk, blokkoló oldattal (1% BSA vagy kecskeszérum 0,1% PBST-ben) szobahőmérsékleten 1 órán át vagy 4 °C-on egy éjszakán át. A diákat ezután PBST-vel háromszor, egyenként 10 percig szobahőmérsékleten öblítettük le, majd 100 uL AF 488-as címkével ellátott kecske anti-egér IgG-vel (Invitrogen, Carlsbad, CA, USA, cat. no. A-11001) szobahőmérsékleten 1 órán át blokkoló oldattal hígítva. A csúszdákat ezután háromszor szobahőmérsékleten 10 percig öblítettük PBS-sel, és szerelőközeggel fedőlemezekkel borítottuk be őket. A kolokalizációs elemzésekhez a képeket az ImageJ (NIH, Bethesda, MD, USA) JACoP beépülő moduljával elemeztük [18].®
4.13. TNF-α egérszövet-koncentrációk mérése
A szövetmintákat lízispufferben homogenizáltuk (kb. 79216, Qiagen, Düsseldorf, Németország) 1% NP-40/PBS-ben egy Complete Mini EDTA-mentes proteáz inhibitor koktélban (kb. 11836170001, Roche, Bázel, Svájc), a szöveti lizátumokat pedig egér TNF-alfa Quantikine ELISA készlettel (kat. sz.) elemeztük egy egér TNF-alfa kvantikin ELISA készlettel (kat. sz. sz. MTA00B, K+ D Systems, Minneapolis, MN, USA) a gyártó utasításai szerint.
4.14. Statisztikai elemzés
Az eredményeket az átlag (SEM) átlagának + standard hibája formájában fejezzük ki. A kísérleti csoportok közötti különbségeket a variancia egyirányú elemzésével (ANOVA) értékeltük. A p < 0,05 értékeket szignifikánsnak fogadtuk el [20,54].
Kiegészítő anyagok
A következő támogató információk a következő címen tölthetők le: https://www.mdpi.com/article/10.3390/ijms23147609/s1.
Szerző közreműködése
Koncepcionálás, T.L., A.H. és L.S.; módszertan, A.H., G.M., J.B., R.P., T.F.P., A.L., A.P.-S., C.V., L.S. és T.L.; validálás, A.H., G.M., J.B., R.P., T.F.P., A.L., A.P.-S., C.V., L.S. és T.L.; formális elemzés, A.H., G.M., J.B., R.P., T.F.P., A.L., C.V., L.S. és T.L.; vizsgálat, A.H., G.M., J.B., R.P., T.F.P., A.L., A.P.-S., C.V., L.S. és T.L.; adatgondozás, A.H. és T.L.; írás — eredeti tervezet készítése, T. L. és A.H.; írás – áttekintés és szerkesztés, A.H., A.L. és T.L.; vizualizáció, A.H., G.M., J.B., R.P., T.F.P. és T.L.; felügyelet, T.L.; projektadminisztráció, T.L. és A.H.; finanszírozási akvizíció, T.L. Minden szerző elolvasta és elfogadta a kézirat megjelent változatát.
Finanszírozás
Az A.H., L.S. és T.L. az 807015. számú támogatási megállapodás alapján finanszírozásban részesült az Innovatív Gyógyszerek Kutatására irányuló Kezdeményezést megvalósító 2 közös vállalkozástól. Ez a közös vállalkozás támogatást kap az Európai Unió Horizont 2020 kutatási és innovációs programjától és az EFPIA-tól. Az A.H., az L.S. és a T.L. az Európai Unió Horizont 2020 kutatási és innovációs programjából is részesült finanszírozásban a Future and Emerging Technologies 863214. számú támogatási megállapodás keretében. Az A.H., L.S. és T.L. támogatást a 2017-2.3.6-TÉT-CN-2018-00023 számú támogatás is támogatta. Az A.H., L.S., A.L., P.S.A., C.V. és T.L. támogatást a Nemzeti Kutatási, Fejlesztési és Innovációs Hivatal 2020-1.1.6-JÖVŐ-2021-00012 számú támogatásával támogatták.
Intézményi Felülvizsgálati Testületi Nyilatkozat
Minden állatkísérletet a nemzeti és intézményi etikai irányelvek szerint végeztünk. Az állatkísérleti vizsgálati protokollt a Biológiai Kutatóközpont Intézményi Állatetikai Bizottsága (BRC-MÁB/2020-3 protokollkód, amelyet 2020. március 12-én hagytak jóvá) és az Állatkísérletekkel Foglalkozó Nemzeti Tudományos Etikai Bizottság (XVI./719/2020. sz. protokollkód, amelyet 2020. május 8-án hagytak jóvá) hagyta jóvá.
Tájékoztatáson alapuló beleegyező nyilatkozat
Nem alkalmazható.
Adat-rendelkezésre állási nyilatkozat
Az adatokat az árucikk vagy a kiegészítő anyagok tartalmazzák.
Összeférhetetlenség
A szerzők nem nyilatkoznak összeférhetetlenségről.
Hivatkozások
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H. J.; Hsueh, P.R. Súlyos akut légzőszervi szindróma koronavírus 2 (SARS-CoV-2) és koronavírus-betegség-2019 (COVID-19): A járvány és a kihívások. Int. J. Antimikrob. Ügynökök 2020, 55, 105924. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Israfil, S.M.H.; Sarker, M.M.R.; Rashid, P.T.; Talukder, A.A.; Kawsar, K.A.; Kán, F.; Akhter, S.; Poh, C.L.; Mohamed, I.N.; Ming, L.C. A COVID-19 klinikai jellemzői és diagnosztikai kihívásai: Frissítés globális perspektívából. Eleje. Közegészségügy 2020, 8, 567395. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Goyal, M.; Tewatia, N.; Vashisht, H.; Dzsain, R.; Kumar, S. Új koronavírus (COVID-19); Globális erőfeszítések és hatékony vizsgálati gyógyszerek: áttekintés. J. Fertőz. Közegészségügy 2021, 14, 910–921. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Wen, W.; Csen, C.; Tang, J.; Wang, C.; Zhou, M.; Cseng, Y.; Zhou, X.; Wu, Q.; Zhang, X.; Feng, Z.; és mtsai. Három új orális antivirális antivirális kezelés (molnupiravir, fluvoxamin és Paxlovid) hatékonysága és biztonságossága a COVID-19 kezelésére: metaanalízis. Ann. Med. 2022, 54, 516–523. [Google Tudós] [Keresztref]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. A SARS-CoV-2 sejtbevitel az ACE2-től és a TMPRSS2-től függ, és egy klinikailag bizonyított proteáz inhibitor blokkolja. Sejt 2020, 181, 271–280. [Google Tudós] [Keresztref]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M. G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: A SARS-CoV-2 fő sejtbevezetési receptora. Tüdő 2020, 198, 867–877. [Google Tudós] [Keresztref]
- Shen, X.R.; Geng, R.; Li, Q.; Csen, Y.; Li, S.F.; Wang, Q.; Min, J.; Jang, Y.; Li, B.; Csiang, R.D.; et al. ACE2-független T limfociták fertőzése SARS-CoV-2-vel. Jelátalakító. Cél. Ther. 2022, 7, 83. [Google Tudós] [Keresztref]
- Karthika, T.; József, J.; Das, V.R.A.; Nair, N.; Charulekha, P.; Roji, M.D.; Raj, V.S. SARS-CoV-2 sejtes belépés független az ACE2 citoplazmatikus domén jelátviteltől. Sejtek 2021, 10, 1814. [Google Tudós] [Keresztref]
- Liu, J.; Lu, F.; Csen, Y.; Eke, E.; Qin, J. Integrin közvetíti a SARS-CoV-2 vírus sejtbelépését az ACE2 sejtreceptortól függetlenül. J. Biol. Chem. 2022, 298, 101710. [Google Tudós] [Keresztref]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor elismerés az új koronavírus által Vuhanból: A SARS koronavírus évtizedes strukturális tanulmányain alapuló elemzés. J. Virol. 2020, 94, e00127-20. [Google Tudós] [Keresztref]
- Shou, S.; Liu, M.; Jang, Y.; Kang, N.; Dal, Y.; Tan, D.; Liu, N.; Wang, F.; Liu, J.; Xie, Y. Állatmodellek a COVID-19-hez: Hörcsögök, egér, vadászgörény, nyérc, favágó és főemlősök. Eleje. Mikrobiol. 2021, 12, 626553. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Schuurs, Z.P.; Hammond, E.; Elli, S.; Rudd, T.R.; Mycroft-West, C.J.; Lima, M.A.; Skidmore, M.A.; Karlsson, R.; Csen, Y.H.; Bagdonaite, I.; és mtsai. Feltételezett glikozaminoglikán kötőhely bizonyítéka a glikozilált SARS-CoV-2 tüskefehérje N-terminális doménénénén. Számítás. Struct. Biotechnológia. Üdv. 2021, 19, 2806–2818. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Kim, S.Y.; Dzsin, W.; Sood, A.; Montgomery, D.W.; Grant, O.C.; Fuster, M.M.; Fu, L.; Dordick, J.S.; Woods, R. J.; Zhang, F.; és mtsai. A heparin és a súlyos akut légzőszervi szindrómával kapcsolatos koronavírus 2 (SARS-CoV-2) tüske glikoprotein-kötő kölcsönhatások jellemzése. Vírusellenes Res. 2020, 181, 104873. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Clausen, T. M.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Festő, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. A SARS-CoV-2 fertőzés a sejtes heparán-szulfáttól és az ACE2-től függ. Sejt 2020, 183, 1043–1057. [Google Tudós] [Kereszthivatkozás] [PubMed]
- De Pasquale, V.; Quiccione, M.S.; Tafuri, S.; Avallone, L.; Pavone, L.M. Heparán-szulfát proteoglikánok vírusfertőzésben és kezelésben: Különös hangsúly a SARS-CoV-2-re. Int. J. Mol. Sci. 2021, 22, 6574. [VISSZA] [Google Tudós] [Keresztref]
- Paiardi, G.; Richter, S.; Oreste, P.; Urbinati, C.; Rusnati, M.; Wade, R.C. A heparin kötődése a tüskés glikoproteinhez három mechanizmussal gátolja a SARS-CoV-2 fertőzést. J. Biol. Chem. 2022, 298, 101507. [Google Tudós] [Keresztref]
- Tumova, S.; Woods, A.; Couchman, J.R. Heparan szulfát láncok glipicánból és szindekánokból hasonlóan kötik össze a fibronektin Hep II doménjét a kisebb szerkezeti különbségek ellenére. J. Biol. Chem. 2000, 275, 9410–9417. [Google Tudós] [Keresztref]
- Hudak, A.; Letoha, A.; Szilak L.; Letoha, T. A szindekánok hozzájárulása a SARS-CoV-2 sejtes belépéséhez. Int. J. Mol. Sci. 2021, 22, 5336. [VISSZA] [Google Tudós] [Keresztref]
- Letoha, T.; Keller-Pinter, A.; Kusz, E.; Kolozsi, C.; Bozso, Z.; Tóth, G.; Vizler, C.; Oláh, Z.; Szilak, L. Sejt-áthatoló peptid kihasználta a szindekánokat. Biochim. Biofizs. Acta 2010, 1798, 2258–2265. [Google Tudós] [Keresztref]
- Hudak, A.; Kusz, E.; Domonkos, I.; Josvay, K.; Kodamullil, A.T.; Szilak L.; Hofmann-Apitius, M.; Letoha, T. A szindekánok hozzájárulása az alfa-szinuklein és a tau sejtfelvételéhez és fibrillációjához. Sci. Rep. 2019, 9, 16543. [Google Tudós] [Keresztref]
- Fuki, I.V.; Meyer, M.E.; Williams, K.J. A szindekán transzmembrán és citoplazmatikus doménjei többlépcsős endocitásos útvonalat közvetítenek mosószerrel oldhatatlan membrántutajokkal. Biokémia. J. 2000, 351 Pt 3, 607–612. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Tkachenko, E.; Rodosz, J.M.; Simons, M. Syndecans: Új gyerekek a jelzőblokkban. Kör alakú. 2005, 96, 488–500. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Couchman, J.R.; Gopal, S.; Lim, H.C.; Norgaard, S.; Multhaupt, H.A. Fell-Muir Előadás: Syndecans: A perifériás coreceptoroktól a sejtmag viselkedésének mainstream szabályozóiig. Int. J. Exp. Pathol. 2015, 96, 1–10. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Hudak, A.; Veres, G.; Letoha, A.; Szilak L.; Letoha, T. Syndecan-4 a SARS-CoV-2 delta variáns kiváló sebességváltójának kulcsfontosságú segítője. Int. J. Mol. Sci. 2022, 23, 796. [VISSZA] [Google Tudós] [Keresztref]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparán-szulfát proteoglikánok. Hideg tavaszi harb. Perspektíva. Biol. 2011, 3, a004952. [Google Tudós] [Keresztref]
- Capila, I.; Linhardt, R.J. Heparin-fehérje kölcsönhatások. Angew. Chem. Int. Szerk. Engl. 2002, 41, 391–412. [Google Tudós] [Keresztref]
- Hileman, R.E.; Fromm, J.R.; Weiler, J.M.; Linhardt, R.J. Glikosaminoglikán-fehérje kölcsönhatások: A glikozaminoglikán kötőfehérjék konszenzusos helyeinek meghatározása. Bioessays 1998, 20, 156–167. [Google Tudós] [Keresztref]
- Vallet, S.D.; Klerc, O.; Ricard-Blum, S. Glikosaminoglikán-fehérje kölcsönhatások: A glikozaminoglikán kölcsönhatások első huzata. J. Histochem. Citokémia. 2021, 69, 93–104. [Google Tudós] [Keresztref]
- Raman, R.; Sasisekharan, V.; Sasisekharan, R. Szerkezeti betekintés a fehérje-glikozaminoglikán kölcsönhatások biológiai szerepébe. Chem. Biol. 2005, 12, 267–277. [Google Tudós] [Keresztref]
- Simon Davis, D.A.; Plébánia, C.R. Heparán-szulfát: Mindenütt jelen lévő glikozaminoglikán, több szerepet tölt be az immunitásban. Eleje. Immunol. 2013, 4, 470. [VISSZA] [Google Tudós] [Keresztref]
- Cardin, Kr.u.; Weintraub, H.J. Fehérje-glikozaminoglikán kölcsönhatások molekuláris modellezése. Arterioszklerózis 1989, 9, 21–32. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Mali, M.; Jaakkola, P.; Arvilommi, A.M.; Jalkanen, M. Az emberi szindekán szekvenciája az integrált membrán proteoglikánok új géncsaládját jelzi. J. Biol. Chem. 1990, 265, 6884–6889. [Google Tudós] [Keresztref]
- Billings, P.C.; Pacifici, M. A jelzőfehérjék, növekedési faktorok és más fehérjék kölcsönhatásai heparán-szulfáttal: mechanizmusok és rejtélyek. Összeköt. Szöveti res. 2015, 56, 272–280. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Christianson, H.C.; Övezés, M. Heparan szulfát proteoglikán, mint sejtfelszíni endocitózis receptor. Mátrix Biol. 2014, 35, 51–55. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Letoha, T.; Hudak, A.; Kusz, E.; Pettko-Szandtner, A.; Domonkos, I.; Josvay, K.; Hofmann-Apitius, M.; Szilak, L. A szindekánok hozzájárulása a sejtek internalizálásához és az amiloid-béta fibrillációjához(1-42). Sci. Rep. 2019, 9, 1393. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Parolini, I.; Sargiacomo, M.; Galbiati, F.; Rizzo, G.; Grignani, F.; Engelman, J.A.; Okamoto, T.; Ikezu, T.; Scherer, P.E.; Móra, R.; és mtsai. A caveolin-1 kifejeződése szükséges a caveolin-2 plazmamembránba történő szállításához. A caveolin-2 megtartása a golgi komplex szintjén. J. Biol. Chem. 1999, 274, 25718–25725. [Google Tudós] [Keresztref]
- Saphire, A.C.; Bobardt, M.D.; Zhang, Z.; Dávid, G.; Gallay, P.A. A syndekánok kötődési receptorként szolgálnak az 1-es típusú humán immunhiányos vírushoz a makrofágokon. J. Virol. 2001, 75, 9187–9200. [Google Tudós] [Keresztref]
- Steinfeld, R.; Van Den Berghe, H.; David, G. A fibroblaszt növekedési faktor receptor-1 kihasználtságának és jelátvitelének stimulálása sejtfelszínhez kapcsolódó szindekánokkal és glicénnel. J. Sejt biol. 1996, 133, 405–416. [Google Tudós] [Keresztref]
- Uhlen, M.; Bjorling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; és mtsai. Emberi fehérje atlasz normál és rákos szövetekhez antitest-proteomika alapján. Mol. Sejtproteomika 2005, 4, 1920–1932. [Google Tudós] [Keresztref]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Az emberi proteom szövetalapú térképe. Tudomány 2015, 347, 1260419. [Google Tudós] [Keresztref]
- Sjostedt, E.; Zhong, W.; Fagerberg, L.; Karlsson, M.; Mitsios, N.; Adori, C.; Oksvold, P.; Edfors, F.; Limiszewska, A.; Hikmet, F.; és mtsai. Az emberi, sertés- és egéragyban lévő fehérjekódoló gének atlasza. Tudomány 2020, 367, eaay5947. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Maggi, E.; Canonica, G.W.; Moretta, L. COVID-19: Megválaszolatlan kérdések az immunválaszról és a patogenezisről. J. Allergia Clin. Immunol. 2020, 146, 18–22. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Sharun, K.; Dhama, K.; Pawde, A.M.; Gortazar, C.; Tiwári, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J.; de la Fuente, J.; Michalak, I.; Attia, Y.A. SARS-CoV-2 állatokban: Ismeretlen tározógazdák lehetősége és közegészségügyi következmények. Állatorvos 2021. negyedév, 41. szám, 181–201. [Google Tudós] [Keresztref]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. A SARS-CoV-2 sejtekbe való belépésének mechanizmusai. Mol. Ford. Sejt biol. 2022, 23, 3–20. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Colombo, V.C.; Sluydts, V.; Marien, J.; Vanden Broecke, B.; Van Houtte, N.; Leirs, W.; Jacobs, L.; Iserbyt, A.; Hubert, M.; Heyndrickx, L.; et al. SARS-CoV-2 megfigyelés norvég patkányokban (Rattus norvegicus) Antwerpen csatornarendszeréből, Belgiumból. Transzbound. Jelenik meg. Dis. 2021. [Google Tudós] [CrossRef] [PubMed]
- Bartlett, A.H.; Park, P.W. Heparán-szulfát proteoglikánok fertőzésben. A Glycans-ban a betegségekben és a terápiákban; Pavão, M.S.G., szerk.; Springer: Berlin/Heidelberg, Németország, 2011; 31–62. oldal. [Google Tudós]
- Cagno, V.; Tseligka, E.D.; Jones, S.T.; Tapparel, C. Heparan-szulfát proteoglikánok és vírusmegkötők: Valódi receptorok vagy adaptációs torzítás? Vírusok 2019, 11, 596. [Google Tudós] [Keresztref]
- Bobardt, M.D.; Lazac, P.; Wang, L.; Esko, J.D.; Gabuzda, D.; Fiala, M.; Trono, D.; Van der Schueren, B.; Dávid, G.; Gallay, P.A. A proteoglikánok hozzájárulása az emberi immunhiányos vírus 1. típusú agyi inváziójához. J. Virol. 2004, 78, 6567–6584. [Google Tudós] [Keresztref]
- Pizzato, M.; Baraldi, C.; Boscato Sopetto, G.; Finozzi, D.; Pogány, C.; Nemzsidó, M. D.; Marconi, R.; Paladino, D.; Raoss, A.; Riedmiller, I.; et al. SARS-CoV-2 és a gazdasejt: Mese az interakciókról. Eleje. Virol. 2022, 1, 815388. [Google Tudós] [Keresztref]
- Gadanec, L.K.; McSweeney, K.R.; Qaradakhi, T.; Ali, B.; Zulli, A.; Apostolopoulos, V. Használhat-e a SARS-CoV-2 vírus több receptort a gazdasejtekbe való belépéshez? Int. J. Mol. Sci. 2021, 22, 992. [VISSZA] [Google Tudós] [Keresztref]
- Bayati, A.; Kumar, R.; Ferenc, V.; McPherson, P.S. A SARS-CoV-2 a vírus bejutása után clathrin által közvetített endocitózissal fertőzi meg a sejteket. J. Biol. Chem. 2021, 296, 100306. [Google Tudós] [Keresztref]
- Ghosh, S.; Dellibovi-Ragheb, T.A.; Kerviel, A.; Pak, E.; Qiu, Q.; Fisher, M.; Takvorian, P.M.; Bleck, C.; Hsu, V.W.; Fehr, A.R.; et al. béta-koronavírusok lizoszómákat használnak a kilépéshez a bioszintézis szekréciós útvonal helyett. Sejt 2020, 183, 1520–1535. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Prydz, K.; Saraste, J. A koronavírusok életciklusa és rejtélyes kijutása. Mol. Mikrobiol. 2022, 117, 1308–1316. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Cesar-Silva, D.; Pereira-Dutra, F.S.; Moraes Giannini, A.L.; Jacques, G.d.A.C. Az endolizomoszóma-rendszer: A SARS-CoV-2 savtesztje. Int. J. Mol. Sci. 2022, 23, 4576. [VISSZA] [Google Tudós] [Kereszthivatkozás] [PubMed]
- Bareford, L.M.; Swaan, P.W. Endocytikus mechanizmusok a célzott gyógyszeradagoláshoz. Adv. Drug Deliv. Rev. 2007, 59, 748–758. [Google Tudós] [Kereszthivatkozás] [PubMed]
- Zimmermann, P.; Tomatis, D.; Rosas, M.; Grootjans, J.; Leenaerts, I.; Degeest, G.; Reekmans, G.; Coomans, C.; David, G. A syntenin, egy szindekánkötő PDZ fehérje jellemzése a sejtadhéziós helyek és mikroszálak összetevőjeként. Mol. Biol. Cell 2001, 12, 339–350. [Google Tudós] [Keresztref]
- Chen, K.; Williams, K.J. Molekuláris mediátorok a syndecan-1 tutajfüggő endocitózisához, amely egy erősen konzervált, multifunkcionális receptor. J. Biol. Chem. 2013, 288, 13988–13999. [Google Tudós] [Keresztref]
- Karnovszkij, M. J.; Karnovszkij, M. J.; Karnovszkij, M. J.; Karnovszkij, M.L.; Karnovszkij, M.J. Magas ozmolalitású formaldehid-glutaraldehid fixálószer elektronmikroszkópiában való felhasználásra. J. Sejt biol. 1965. 27., 137. [VISSZA] [Google Tudós]
- Zsiros, O.; Nagy G.; Patai, R.; Solymosi, K.; Gasser, U.; Polgar, T. F.; Garab, G.; Kovács L.; Horcsik, Z.T. Hasonlóságok és különbségek a kadmium és a króm toxikus koncentrációjának a Chlorella variabilis tylakoid membránjainak szerkezetére és funkcióira gyakorolt hatásában. Eleje. Növény Sci. 2020, 11, 1006. [VISSZA] [Google Tudós] [Keresztref]
- Millonig, G. Módosított eljárás vékony szakaszok ólomfestésére. J. Biofis. Biokémia. Cytol. 1961, 11, 736–739. [Google Tudós] [Keresztref]
- Nakase, I.; Niwa, M.; Takeuchi, T.; Sonomura, K.; Kawabata, N.; Koike, Y.; Takehashi, M.; Tanaka, S.; Ueda, K.; Simpson, J.C.; és mtsai. Az argininben gazdag peptidek sejtfelvétele: A makropinocitózis és az aktin átrendeződés szerepe. Mol. Ther. 2004, 10, 1011–1022. [Google Tudós] [Keresztref]
- Nakase, I.; Tadokoro, A.; Kawabata, N.; Takeuchi, T.; Katoh, H.; Hiramoto, K.; Negishi, M.; Nomizu, M.; Sugiura, Y.; Futaki, S. Az argininben gazdag peptidek és a membránnal összefüggő proteoglikánok kölcsönhatása döntő fontosságú az aktin szervezet és a makropinocitózis indukálásához. Biokémia 2007, 46, 492–501. [Google Tudós] [Kereszthivatkozás] [PubMed]
A kiadó megjegyzése: Az MDPI semleges marad a közzétett térképeken és intézményi kapcsolatokban szereplő joghatósági követelések tekintetében. |
© 2022 a szerzőktől. MdPI engedélyes, Bázel, Svájc. Ez a cikk egy nyílt hozzáférésű cikk, amelyet a Creative Commons Attribution (CC BY) licenc (https://creativecommons.org/licenses/by/4.0/) feltételei szerint terjesztenek.